

In vitro spontaneous differentiation of human breast cancer stem cells and methods to control this process
Copyrights: Phuc Van Pham, Binh Thanh Vu, Nhan Lu-Chinh Phan, Hanh Thi Le, Ngoc Kim Phan, 2015. License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Abstract
Breast cancer stem cells were considered as origins of breast cancer. Previously published studies showed that breast cancer stem cells exhibited high multi-drug resistance. This study aimed to evaluate the spontaneous differentiation of human breast cancer stem cells and investigate some in vitro conditions to control this process. Human breast cancer stem cells (BCSCs) were sorted from primary culture of breast malignant tumors based on expression of CD44 and CD24. The in vitro spontaneous differentiation of BCSCs was evaluated in the popular culture medium DMEM/F12 supplemented with 10% fetal bovine serum (FBS), 1% antibiotic-antimycotic. There were some different methods to control the spontaneous differentiation of BCSCs included free serum culture, mammosphere culture, basic fibroblast growth factor and epidermal growth factor supplement to serum medium, and hypoxia culture. The results showed that BCSCs always were spontaneously differentiated in vitro in the popular culture medium DMEM/F12 plus 10% FBS. The percentage of BCSCs gradually decreased according to sub-culture times and became stable after 20 sub-culture times. All investigated methods could not completely inhibit the spontaneous differentiation of BCSCs. Serum-free culture combined with hypoxia condition had strongest inhibition of this process. These results demonstrated that the spontaneous differentiation is nature process of BCSCs; therefore this process should be determined and suitably controlled depending on different experiments.
Introduction
Breast tumors are thought to originate from breast cancer stem cells (BCSCs) that were first discovered by Al-Hajj and colleagues Al-Hajj et al., 2003. Because BCSCs have been shown to be multi-drug resistant, radiation-resistant, and involved in metastasis and cancer relapse Al-Ejeh et al., 2011Liu et al., 2010Liu and Wicha, 2010Xie et al., 2012, they are considered to be the target for all new breast cancer therapies Ginestier et al., 2010Liu and Wicha, 2010. Some preclinical studies and clinical trials that have targeted BCSCs have had promising results Ahmadipour et al., 2015Bostad et al., 2015Gupta et al., 2009Hu et al., 2015Jiang et al., 2015Kai et al., 2015Luo et al., 2015Montales et al., 2012Wu et al., 2015Zhang et al., 2014. The data confirm that BCSCs play a critical role in the initiation oftumors. Therefore, BCSCs are an important and valuable model for drug discovery as well as for the development of novel therapies.
As of 2003, BCSCs have been isolated using different methods by several groups. Culturing BCSCs, as described in previous studies, has involved the use of various culture media and techniques, including the use of standard DMEM/F12 medium supplemented with 10% fetal bovine serum (FBS) for adherent cultures Pham et al.,2011Van Phuc et al., 2011, serum-reduced medium for mammospheres Carmody et al., 2012 Gupta et al., 2011, and more recently, serum-free medium to maintain BCSCs in culture de la Mare et al., 2013Grimshaw et al., 2008Lombardo et al.,2015Manuel Iglesias et al., 2013Montales et al., 2012Wang et al., 2014.
In some recent publications, oxygen concentration has been shown to also affect drug resistance Crowder et al., 2014Samanta et al., 2014 and stemness of BCSCs Mimeault and Batra, 2013van den Beucken et al., 2014. Therefore, in this study we investigated the effects of components of media (FBS and growth factors) and oxygen concentration on spontaneous differentiation of BCSCs in vitro. Our results suggest that there are suitable cell culture conditions in which to maintain the stemness of BCSCs.
Material and Methods
Isolation of BCSCs
BCSCs were isolated according to previously published research Pham et al., 2011 Van Phuc et al., 2011. CD44+CD24-BCSCs were sorted from a breast cancer cell population using flow cytometry (FASC-jazz, BD Bioscience). Briefly, BCSCs were stained with monoclonal antibodies CD44-FITC and CD24-APC. CD44+CD24- BCSCs were sorted and re-confirmed using a FACSCalibur flow cytometer. CD44+CD24-BCSCs with 100% purity were used in further experiments.
BCSC culture
In this study, three kinds of media were used: DMEM/F12 supplemented with 10% FBS; DMEM/F12 supplemented with 10% FBS, with 10 ng/ml EGF and 10 ng/ml FGF; and mammosphere medium ( Table 1 ). In addition, two oxygen concentrations, 10 and 21%, were tested. Six groups, made up of a combination of three kinds of medium and two oxygen concentrations, were investigated as shown in Table 1 . BCSCs were cultured in 96-multi-well plates at 104 cells/well. All plates were cultured in a tri-gas incubator at 37°C and 5% CO2.
Flow cytometry analysis
Cells were washed twice in phosphate-buffered saline containing 1% bovine serum albumin (Sigma-Aldrich, St Louis, MO). Fc receptors were blocked by incubation with immunoglobulin G (Santa Cruz Biotechnology, CA) on ice for 15 min. Cells were stained with anti-CD44-FITC and anti-CD24-APC monoclonal antibodies (BD Biosciences, Franklin Lakes, NJ, USA) at 4° C for 30 min. After washing, cells were analyzed using a FACSCalibur flow cytometer (BD Biosciences) and CellQuest Pro software (BD Biosciences) using data acquired from 10,000 events.
Doxorubicin resistance assay
BCSCs were cultured as shown in Table 1 and then seeded at a density of 0.4 × 104 cells per well in 24-well plates (Nunc) in DMEMF12/10% FBS. After 24 h, cells were treated with 0, 1, 3, and 6 μg/ml doxorubicin (Sigma-Aldrich, St. Louis, MO, USA) for 48 h. Cells were then subjected to apoptotic analysis.
Annexin V/PI assay
Apoptosis was investigated using flow cytometry with annexin V and propidium iodide (PI; BD Biosciences). Cells that had been grown under the six different culture conditions were washed twice in PBS and fixed in cold 70% ethanol for at least 3 h at 4°C. Subsequently, cells were washed twice in PBS and stained with 1 ml of PI (20 g/ml). A 50 μl volume of RNase A (10 μg/ml) was added to samples and incubated for 3h at 4°C. Stained cells were analyzed using flow cytometry and CellQuest Pro software.
In vivotumorigenesis assay
Female (5-6 weeks old) NOD/severe-combined immunodeficient (SCID) mice (NOD.CB17-Prkdcscid/J; Charles River Laboratories, Wilmington, MA) were subcutaneously injected with 105 cells/mouse, with three mice per group. Mice were visually monitored for tumor formation at 7, 14 and 21 days post-injection. The Institutional Animal Care and Use Committee of Stem Cell Research and Application Laboratory, University of Science, VNU-HCM approved all animal experiments. s
Statistical analysis
All experiments were performed in triplicate. The significance of differences between mean values was assessed using the t-test and ANOVA. A p-value less than 0.05 was considered to be significant. Data were analyzed using Prism 6 software.
Results
Spontaneous differentiation of BCSCs in vitro
The percentage of sorted CD44+CD24-BCSCs gradually decreased after 1, 2, 4 and 8 weeks of culture in DMEM/F12 supplemented with 10% FBS and 1% anti-biotic-antimycotic, although cell morphology was not significantly different. The percent of CD44+CD24-BCSCs decreased from 100% at start to 65.43±15.21%, 47.39±17.31%, 33.87±9.81%, and 12.31+4.52% after 1, 2, 4 and 8 weeks, respectively. Moreover, doxorubicin resistance also dramatically decreased after 4 and 8 weeks of culture. In fact, the percent of apoptotic cells significantly increased after 4 and 8 weeks of culture compared with after 1 and 2 weeks (20.41±43%, 32±9.41%, 54.78±14.76%, and 68.41±14.39% apoptotic cells after 1, 2, 4 and 8 weeks, respectively). The in vivotumorigenesis assay also showed that tumorigenesis of cultured cells decreased after 8 weeks of culture (100% of mice bearing tumors per group after 1, 2 and 4 weeks vs. 75 % mice bearing tumors per group after 8 weeks).
Effects of culture conditions on the CD44+CD24-BCSC population
To optimize culture conditions for BCSCs, a combination of three different culture media and two oxygen concentrations were used. BCSCs were cultured in these six different conditions ( Table 1 ) for 8 weeks. The CD44+CD24- cell population was isolated using flow cytometry and cell shape changes were monitored using an inverted microscope. The results are presented in Figure 1 and showed that cell morphology showed little change between week 1 and week 8 in all groups (see Table 1 ).
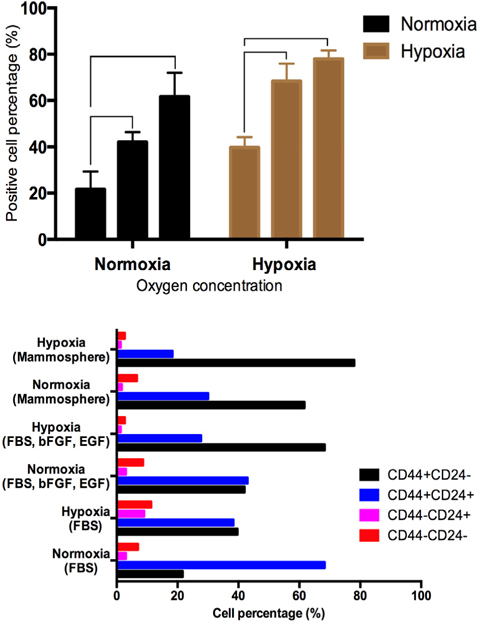
As presented in Figure 1 , the cell population with the BCSC phenotype was different between groups. In the DMEM/F12 supplemented with 10% FBS and a normal level of oxygen, after 4 weeks, there were only 21.57±4.41% CD44+CD24- BCSCs, while this percentage increased when cells were grown in the same medium but under hypoxic conditions (39.57±2.60%). For all media, the hypoxic condition always resulted in a greater percentage of CD44+CD24-BCSCsthan under normoxic conditions (in DMEM/F12 + 10% FBS, 21.67±4.41% vs. 39.67±2.60%; in DMEM/F12 plus FGF and EGF, 42.00±2.52 vs. 68.33±4.41%, and in mammos-phere medium 61.67±6.01% vs. 78.00±2.08%, under normoxic and hypoxic conditions, respectively).
We found that culture medium and oxygen concentration affected the BCSC phenotype in culture, as shown in Figure 1 . Effects of FBS, high oxygen concentrations as well as a lack of FGF and EGF resulted in the differentiation of BCSCs with a CD44+CD24+phenotype. In medium with 10% FBS in DMEM/F12 and under normoxic conditions, there were up to 70% CD44+CD24+ cells after 4 weeks. This result also showed that BCSCs underwent spontaneous differentiation during in vitro culture.
Effect of culture medium on drug resistance of BCSCs
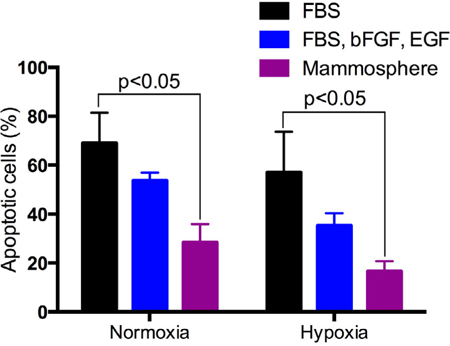
BCSCs cultured under different conditions also exhibited different levels of drug resistance. Fig. 4 shows that BCSCs cultured in DMEM/F12 supplemented with 10% FBS were more sensitive to doxorubicin than BCSCs cultured in DMEM/F12 supplemented with 10% FBS and EGF and bFGF or mammospheres under both normoxic and hypoxic conditions. Under normoxic conditions, doxorubicin treatment of BCSCs cultured in DMEM/F12 plus 10% FBS, DMEM/F12 plus 10% FBS, EGF and FGF, or in mammosphere culture medium resulted in a gradual decrease in the percentage of apoptotic cells from 63.00±8.4 to 44.50±12.96 to 22.50±8.25%, respectively. Similarly, under hypoxic conditions, these percentages were 63.00±8.485, 44.50±12.96, and 22.50±8.250%, respectively. A decrease in the drug sensitive cell population resulted in an increase in the drug resistant cell population ( Figure 2 ).
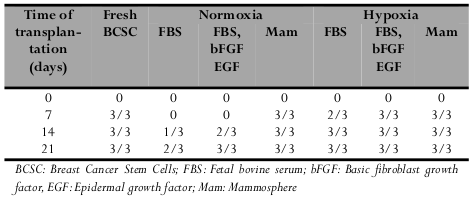
Culture conditions also affected tumorigenesisin vivo. Based on the results shown in Table 2 , BCSCs in mammospheres under both normoxic and hypoxic conditions retained their tumorigenicity when 105 cells were injected into mice, while cells cultured under adherent culture conditions in DMEM/F12 plus 10% FBS or 10% FBS with EGF and FGF showed reduced tumorigenicity ( Table 2 ).
Discussion
Al-Hajj and colleagues discovered BCSCs more than 10 years ago Al-Hajj et al.,2003. BCSCs have since become targets for drugs as well as new therapies, and have become central to breast cancer treatment. Therefore, it is essential to have a ready supply of BCSCs to pursue drug discovery as well as therapy development. However, we show in this study that culture conditions including culture medium and oxygen concentration can affect the stemness of BCSCs during their time in culture.
In the first experiment, the results showed that BCSCs readily differentiated into different types of breast cancer cells. In fact, given the particular characteristics of stem cells, BCSCs can differentiate into specific cells Pham et al., 2011. In this case, BCSCs differentiated into breast cancer cells. The medium containing FBS strongly triggered BCSC differentiation. FBS, which contains a pool of undetermined factors and components, caused the differentiation of BCSCs. Although FBS is an essential component of cell culture medium, it has been shown to cause differentiation of certain kinds of stem cells such as neural and embryonic, as well as germ cells Bettiol et al.,2007Franke et al., 2014Hung and Young, 2006Tateishi et al., 2008Zahir et al.,2009.
In the next experiments, results further showed that in medium supplemented with FBS, a high percentage of BCSCs lost their stemness. This demonstrated that FBS was the differentiating factor. Conversely, BCSCs retained their stemness when cultured in medium lacking FBS. Studies in which proliferation of stem cells was required, therefore, have used serum-free medium.
This study also showed that the oxygen concentration affected maintenance of stemness of BCSCs. A high oxygen concentration triggered the differentiation of BCSCs. In fact, a high concentration of oxygen will increase the amount of radical oxygen species (ROS). ROS also acted as a differentiating factor for stem cells in previous studies. ROS can interfere with certain signaling pathways related to stemness. Hypoxic cultures have been shown in some studies to increase the expression of certain stemness-related genes. Hypoxia-inducible factor (HIF) was found to induce the expression of human embryonic stem cell markers in cancercells Mathieu et al., 2011. Hypoxia has also been shown to prevent etoposide-induced DNA damage in cancercells through a mechanism involving HIF-1 Sullivan and Graham, 2009. More importantly, hypoxia was found to induce miR-424, which decreases tumor sensitivity to chemotherapy by inhibiting apoptosis Zhang et al.,2014. Furthermore, hypoxia could also induce Jagged2, promoting breast cancer metastasis and self-renewal of cancerstem-like cells Xing et al., 2011. Finally, hypoxia was shown to induce expression of HIF-1, which directly enhanced transcriptional activity of stem cell factor (SCF) in response to hypoxia and epidermal growth factor (EGF) Han et al., 2008.
Conclusion
BCSCs can spontaneously differentiate into breast cancer cells. We showed that both the type of culture medium and concentration of oxygen strongly influenced BCSC differentiation. FBS and ROS were also differentiation factors. BCSCs were also found to differentiate into breast cancer cells in medium supplemented with FBS and under normoxic conditions. Therefore, proliferation of BCSCs requires that they be cultured in serum-free medium under hypoxic conditions. These results hoped that there was a significant contribution to BCSC study. However, in this study we showed that it was difficult to culture BCSCs while maintaining 100% stemness (CD44+CD24- BCSC phenotype). Further experiments will need to be performed to improve upon these results.
Abbreviations
BCSCS: Breast cancer stem cells; EGF Epidermal growth factor; FBS: Fetal bovine serum; HIF: Hypoxiainducible factor; SCF: Stem cell factor
References
-
F.
Ahmadipour,
M.I.
Noordin,
S.
Mohan,
A.
Arya,
M.
Paydar,
C.Y.
Looi,
Y.S.
Keong,
E.N.
Siyamak,
S.
Fani,
M.
Firoozi.
Koenimbin, a natural dietary compound of Murraya koenigii (L) Spreng: inhibition of MCF7 breast cancer cells and targeting of derived MCF7 breast cancer stem cells (CD44(+)/CD24(-/low)): an in vitro study. Drug Des Devel Ther.
2015;
9
:
1193-1208
.
-
F.
Al-Ejeh,
C.E.
Smart,
B.J.
Morrison,
G.
Chenevix-Trench,
J.A.
Lopez,
S.R.
Lakhani,
M.P.
Brown,
K.K.
Khanna.
Breast cancer stem cells: treatment resistance and therapeutic opportunities. Carcinogenesis.
2011;
32
:
650-658
.
-
M.
Al-Hajj,
M.S.
Wicha,
A.
Benito-Hernandez,
S.J.
Morrison,
M.F.
Clarke.
Prospective identification of tumorigenic breast cancer cells. Proceedings of the National Academy of Sciences of the United States of America.
2003;
100
:
3983-3988
.
-
E.
Bettiol,
L.
Sartiani,
L.
Chicha,
K.H.
Krause,
E.
Cerbai,
M.E.
Jaconi.
Fetal bovine serum enables cardiac differentiation of human embryonic stem cells. Differentiation.
2007;
75
:
669-681
.
-
M.
Bostad,
C.E.
Olsen,
Q.
Peng,
K.
Berg,
A.
Hogset,
P.K.
Selbo.
Light-controlled endosomal escape of the novel CD133-targeting immunotoxin AC133-saporin by photochemical internalization - A minimally invasive cancer stem cell-targeting strategy. J Control Release.
2015;
206
:
37-48
.
-
L.C.
Carmody,
A.R.
Germain,
L.
VerPlank,
P.P.
Nag,
B.
Munoz,
J.R.
Perez,
M.A.
Palmer.
Phenotypic high-throughput screening elucidates target pathway in breast cancer stem cell-like cells. J Biomol Screen.
2012;
17
:
1204-1210
.
-
S.W
Crowder,
Y.S.
Hwang,
H.J.
Sung.
Cancer Stem Cells under Hypoxia as a Chemoresistance Factor in Breast and Brain. Curr Pathobiol Rep.
2014;
2
:
33-40
.
-
J.A.
Mare,
J.N.
Sterrenberg,
M.G.
Sukhthankar,
M.T.
Chiwakata,
D.R.
Beukes,
G.L.
Blatch,
A.L.
Edkins.
Assessment of potential anti-cancer stem cell activity of marine algal compounds using an in vitro mammosphere assay. Cancer Cell Int.
2013;
13
:
39
.
-
J.
Franke,
V
Abs,
G.
Abraham.
Comparative study of the effects of fetal bovine serum versus horse serum on growth and differentiation of primary equine bronchial fibroblasts. BMC Vet Res.
2014;
10
:
119
.
-
C.
Ginestier,
E.
Charafe-Jauffret,
D.
Birnbaum.
Targeting breast cancer stem cells: fishing season open!. Breast cancer research : BCR.
2010;
12
:
312
.
-
M.J.
Grimshaw,
L.
Cooper,
K.
Papazisis,
J.A.
Coleman,
H.R.
Bohnenkamp,
L.
Chiapero-Stanke,
J.
Taylor-Papadimitriou,
J.M.
Burchell.
Mammosphere culture of metastatic breast cancer cells enriches for tumorigenic breast cancer cells. Breast cancer research : BCR.
2008;
10
:
R52
.
-
P.B.
Gupta,
C.M.
Fillmore,
G.
Jiang,
S.D.
Shapira,
K.
Tao,
C.
Kuperwasser,
E.S.
Lander.
Stochastic state transitions give rise to phenotypic equilibrium in populations of cancer cells. Cell.
2011;
146
:
633-644
.
-
P.B.
Gupta,
T.T.
Onder,
G.
Jiang,
K.
Tao,
C.
Kuperwasser,
R.A.
Weinberg,
E.S.
Lander.
Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell.
2009;
138
:
645-659
.
-
Z.B.
Han,
H.
Ren,
H.
Zhao,
Y.
Chi,
K.
Chen,
B.
Zhou,
Y.J.
Liu,
L.
Zhang,
B.
Xu,
B.
Liu.
Hypoxia-inducible factor (HIF)-1 alpha directly enhances the transcriptional activity of stem cell factor (SCF) in response to hypoxia and epidermal growth factor (EGF). Carcinogenesis.
2008;
29
:
1853-1861
.
-
K.
Hu,
H.
Zhou,
Y.
Liu,
Z.
Liu,
J.
Liu,
J.
Tang,
J.
Li,
J.
Zhang,
W.
Sheng,
Y.
Zhao.
Hyaluronic acid functional amphipathic and redox-responsive polymer particles for the co-delivery of doxorubicin and cyclopamine to eradicate breast cancer cells and cancer stem cells. Nanoscale.
2015;
7
:
8607-8618
.
-
C.H.
Hung,
T.H.
Young.
Differences in the effect on neural stem cells of fetal bovine serum in substrate-coated and soluble form. Biomaterials.
2006;
27
:
5901-5908
.
-
F.
Jiang,
Y.
Li,
J.
Mu,
C.
Hu,
M.
Zhou,
X.
Wang,
L.
Si,
S.
Ning,
Z.
Li.
Glabridin inhibits cancer stem cell-like properties of human breast cancer cells: An epigenetic regulation of miR-148a/SMAd2 signaling. Mol Carcinog.
2015
.
-
M.
Kai,
N.
Kanaya,
S.V.
Wu,
C.
Mendez,
D.
Nguyen,
T.
Luu,
S.
Chen.
Targeting breast cancer stem cells in triple-negative breast cancer using a combination of LBH589 and salinomycin. Breast cancer research and treatment.
2015;
151
:
281-294
.
-
H.
Liu,
M.R.
Patel,
J.A.
Prescher,
A.
Patsialou,
D.
Qian,
J.
Lin,
S.
Wen,
Y.F.
Chang,
M.H.
Bachmann,
Y.
Shimono.
Cancer stem cells from human breast tumors are involved in spontaneous metastases in orthotopic mouse models. Proceedings of the National Academy of Sciences of the United States of America.
2010;
107
:
18115-18120
.
-
S.
Liu,
M.S.
Wicha.
Targeting breast cancer stem cells. Journal of clinical oncology : official journal of the American Society of Clinical Oncology.
2010;
28
:
4006-4012
.
-
Y.
Lombardo,
A.
de Giorgio,
C.R.
Coombes,
J.
Stebbing,
L.
Castellano.
Mammosphere Formation Assay from Human Breast Cancer Tissues and Cell Lines. 2015;
:
e52671
.
-
M.
Luo,
S.G.
Clouthier,
Y.
Deol,
S.
Liu,
S.
Nagrath,
E.
Azizi,
M.S.
Wicha.
Breast cancer stem cells: current advances and clinical implications. Methods Mol Biol.
2015;
1293
:
1-49
.
-
J.
Manuel Iglesias,
I.
Beloqui,
F.
Garcia-Garcia,
O.
Leis,
A.
Vazquez-Martin,
A.
Eguiara,
S.
Cufi,
A.
Pavon,
J.A.
Menendez,
J.
Dopazo.
Mammosphere formation in breast carcinoma cell lines depends upon expression of E-cadherin. PLoS One.
2013;
8
:
e77281
.
-
J.
Mathieu,
Z.
Zhang,
W.
Zhou,
A.J.
Wang,
J.M.
Heddleston,
C.M.
Pinna,
A.
Hubaud,
B.
Stadler,
M.
Choi,
M.
Bar.
HIF induces human embryonic stem cell markers in cancer cells. Cancer research.
2011;
71
:
4640-4652
.
-
M.
Mimeault,
S.K.
Batra.
Hypoxia-inducing factors as master regulators of stemness properties and altered metabolism of cancer- and metastasis-initiating cells. J Cell Mol Med.
2013;
17
:
30-54
.
-
M.T.
Montales,
O.M.
Rahal,
J.
Kang,
T.J.
Rogers,
R.L.
Prior,
X.
Wu,
R.C.
Simmen.
Repression of mammosphere formation of human breast cancer cells by soy isoflavone genistein and blueberry polyphenolic acids suggests diet-mediated targeting of cancer stemlike/progenitor cells. Carcinogenesis.
2012;
33
:
652-660
.
-
P.V
Pham,
N.T.
Nguyen,
N.H.
Truong,
T.T.
Duong,
D.V.
Le,
K.D.
Truong,
N.K.
Phan.
Differentiation of breast cancer stem cells by knockdown of CD44: promising differentiation therapy. Journal of translational medicine.
2011;
9
:
209
.
-
D.
Samanta,
D.M.
Gilkes,
P.
Chaturvedi,
L.
Xiang,
G.L.
Semenza.
Hypoxia-inducible factors are required for chemotherapy resistance of breast cancer stem cells. Proceedings of the National Academy of Sciences of the United States of America.
2014;
111
:
E5429-5438
.
-
R.
Sullivan,
C.H.
Graham.
Hypoxia prevents etoposide-induced DNA damage in cancer cells through a mechanism involving hypoxia-inducible factor 1. Mol Cancer Ther.
2009;
8
:
1702-1713
.
-
K.
Tateishi,
W.
Ando,
C.
Higuchi,
D.A.
Hart,
J.
Hashimoto,
K.
Nakata,
H.
Yoshikawa,
N.
Nakamura.
Comparison of human serum with fetal bovine serum for expansion and differentiation of human synovial MSC: potential feasibility for clinical applications. Cell Transplant.
2008;
17
:
549-557
.
-
T.
Beucken,
E.
Koch,
K.
Chu,
R.
Rupaimoole,
P.
Prickaerts,
M.
Adriaens,
J.W.
Voncken,
A.L.
Harris,
F.M.
Buffa,
S.
Haider.
Hypoxia promotes stem cell phenotypes and poor prognosis through epigenetic regulation of DICER. Nat Commun.
2014;
5
:
5203
.
-
P.
Van Phuc,
P.L.
Nhan,
T.H.
Nhung,
N.T.
Tam,
N.M.
Hoang,
VG.
Tue,
D.T.
Thuy,
P.K.
Ngoc.
Downregulation of CD44 reduces doxorubicin resistance of CD44CD24 breast cancer cells. OncoTrgets and therapy.
2011;
4
:
71-78
.
-
R.
Wang,
Q.
Lv,
W.
Meng,
Q.
Tan,
S.
Zhang,
X.
Mo,
X.
Yang.
Comparison of mammosphere formation from breast cancer cell lines and primary breast tumors. J Thorac Dis.
2014;
6
:
829-837
.
-
C.H.
Wu,
B.H.
Hong,
C.T.
Ho,
G.C.
Yen.
Targeting cancer stem cells in breast cancer: potential anticancer properties of 6-shogaol and pterostilbene. J Agric Food Chem.
2015;
63
:
2432-2441
.
-
G.
Xie,
J.
Zhan,
Y.
Tian,
Y.
Liu,
Z.
Chen,
C.
Ren,
Q.
Sun,
J.
Lian,
L.
Chen,
J.
Ruan.
Mammosphere cells from high-passage MCF7 cell line show variable loss of tumorigenicity and radioresistance. Cancer Lett.
2012;
316
:
53-61
.
-
F.
Xing,
H.
Okuda,
M.
Watabe,
A.
Kobayashi,
S.K.
Pai,
W
Liu,
K.
Fukuda,
S.
Hirota,
T.
Sugai.
Hypoxia-induced Jagged2 promotes breast cancer metastasis and self-renewal of cancer stem-like cells. Oncogene.
2011;
30
:
4075-4086
.
-
T.
Zahir,
Y.F.
Chen,
J.F.
MacDonald,
N.
Leipzig,
C.H.
Tator,
M.S.
Shoichet.
Neural stem/progenitor cells differentiate in vitro to neurons by the combined action of dibutyryl cAMP and interferongamma. Stem Cells Dev.
2009;
18
:
1423-1432
.
-
D.
Zhang,
Z.
Shi,
M.
Li,
J.
Mi.
Hypoxia-induced miR-424 decreases tumor sensitivity to chemotherapy by inhibiting apoptosis. Cell Death Dis.
2014;
5
:
e1301
.
Comments

Downloads
Article Details
Volume & Issue : Vol 2 No 06 (2015)
Page No.: 290-296
Published on: 2015-06-13
Citations
Copyrights & License
This work is licensed under a Creative Commons Attribution 4.0 International License.
Search Panel
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Google Scholar
Pubmed
Search for this article in:
Google Scholar
Researchgate
- HTML viewed - 6082 times
- Download PDF downloaded - 1649 times
- View Article downloaded - 7 times

{kind=link}
{kind=link}
{kind=link}
{kind=link}