
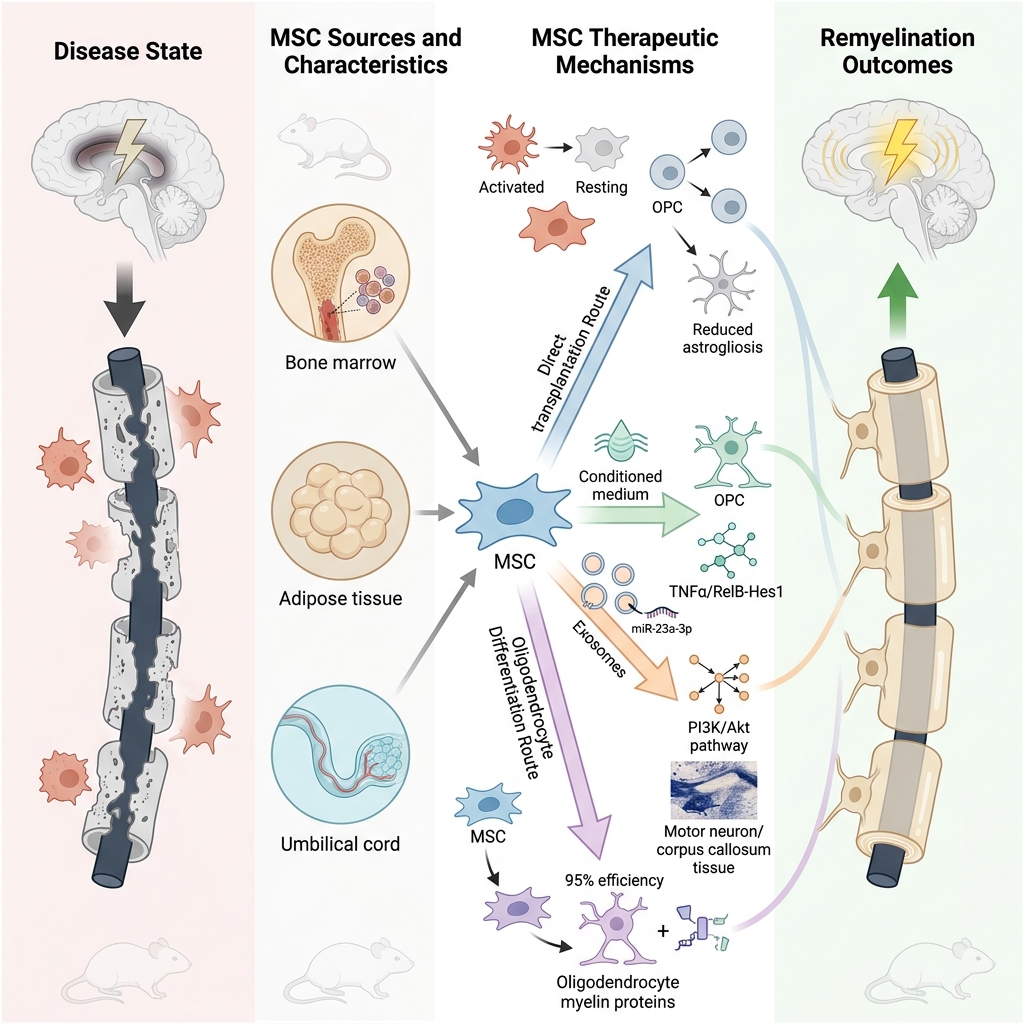

Mesenchymal Stem Cell-Based Therapies for Central Nervous System Demyelination: A Review of Nonhuman Models
-
Universiti Tunku Abdul Rahman Sungai Long Campus

-
Binzhou Medical University
Abstract
Background: Demyelinating diseases of the central nervous system (CNS), such as multiple sclerosis (MS), are characterized by the progressive loss of myelin, leading to neurological deficits. Current therapies demonstrate limited efficacy in promoting remyelination. This review evaluates the therapeutic potential of mesenchymal stem cells (MSCs) and their derivatives for promoting remyelination in nonhuman models of CNS demyelination.
Methods: A comprehensive literature search was conducted across PubMed, Scopus, and Web of Science for studies published between 2020 and 2024. Inclusion criteria focused on in vivo and in vitro studies utilizing MSCs, MSC-conditioned media (MSC-CM), or extracellular vesicles (EVs) within experimental models of CNS demyelination.
Results: Twenty-four studies met the inclusion criteria. The majority of these investigations demonstrated that MSCs enhance oligodendrocyte progenitor cell (OPC) proliferation and differentiation into mature oligodendrocytes (OLs). Key markers evaluated included NG2, PDGFRα, Olig2, and MBP. Furthermore, MSC-CM and EVs exhibited significant neuroprotective and immunomodulatory effects.
Conclusion: MSC-based therapies exhibit promising regenerative potential within preclinical models of CNS demyelination. These findings support further translational research and the advancement of clinical trials targeting remyelination strategies.
Introduction
Demyelinating diseases of the central nervous system (CNS) are characterized by damage to the myelin sheath, a critical structure that ensures efficient nerve signal transmission. Disruption of this sheath impairs neural communication and leads to progressive neurological dysfunction 1. Among these disorders, multiple sclerosis (MS) is the most prevalent, affecting approximately 2.8 million individuals worldwide 2. Typically diagnosed in early adulthood, MS manifests with diverse symptoms including muscle weakness, sensory disturbances, visual impairment, and coordination deficits that profoundly reduce quality of life 3.
Current therapeutic strategies for MS, such as corticosteroids, biologics, disease-modifying therapies (DMTs), and rehabilitation, primarily aim to reduce inflammation and slow disease progression4,5,6,7,8,9. However, none of these interventions directly promote remyelination or neural repair, underscoring a critical gap in treatment and driving interest in regenerative approaches 10. Stem cell-based therapies have emerged as promising candidates for CNS regeneration due to their capacity to modulate immune responses, support neuroprotection, and enhance remyelination 11.
Embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) demonstrate considerable potential given their ability to differentiate into diverse cell types. Nevertheless, their clinical application remains constrained by ethical concerns, safety risks, and high production costs 12,13. Autologous hematopoietic stem cell transplantation (AHSCT) has shown efficacy in inducing remission in aggressive MS cases, but its broader use is limited by treatment-related morbidity, conditioning toxicity, prolonged hospitalization, and unsuitability for progressive MS 14,15.
In contrast, mesenchymal stem cells (MSCs) derived from bone marrow, adipose tissue, or umbilical cord represent a safer and more practical alternative. These cells are readily obtainable, ethically acceptable, and exhibit low immunogenicity, making them suitable for allogeneic therapy. MSCs exert immunomodulatory, neuroprotective, and pro-remyelinating effects through mechanisms such as T-cell regulation and secretion of trophic factors. Clinical trials conducted up to 2025 have reported reduced relapse rates and functional stabilization in patients receiving MSC-based interventions 16,17,18. Notably, umbilical cord-derived MSCs (UC-MSCs) demonstrate high proliferative capacity, while adipose-derived MSCs (AD-MSCs) offer minimally invasive harvesting and high cell yield. MSCs also promote remyelination by enhancing oligodendrocyte progenitor cell (OPC) proliferation, modulating microglial activity, and protecting neural progenitors within neurogenic niches 19. Furthermore, MSC-derived products including conditioned media (MSC-CM) and extracellular vesicles (EVs) have shown significant therapeutic potential in preclinical models 20.
This systematic review synthesizes studies published between 1 January 2020 and 30 November 2025 on MSC-based therapies for remyelination in nonhuman CNS demyelination models. It evaluates outcomes such as OPC activation, oligodendrocyte maturation, and functional recovery, aiming to provide insights that may guide future translational research and clinical applications.
Methods
Literature Search Strategy
A comprehensive literature search was conducted to identify original research articles examining the role of mesenchymal stem cells (MSCs) in promoting remyelination in demyelinating diseases of the central nervous system (CNS). Searches were performed in PubMed, Scopus, and Google Scholar, covering studies published between 1 January 2020 and 30 November 2025. The final search was completed on 30 November 2025. The search strategy combined keywords and Boolean operators: (“Mesenchymal Stem Cells” OR “MSCs”) AND (“Demyelinating Diseases” OR “Multiple Sclerosis”) AND (“Remyelination” OR “Oligodendrocytes”). Only articles published in English were included. The review followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines 21.
Selection of Research Articles/Eligibility Criteria
Studies were selected according to predefined inclusion and exclusion criteria.
Inclusion Criteria
-
Original research articles investigating MSCs or MSC-derived products (e.g., conditioned media [MSC-CM], extracellular vesicles [EVs]) in CNS demyelination.
-
Studies conducted in nonhuman models (in vivo or in vitro).
-
Studies focusing on MSC types such as umbilical cord-derived MSCs (UCMSCs), bone marrow-derived MSCs (BMSCs), adipose-derived MSCs (ADSCs), dental pulp stem cells (DPSCs), and olfactory mesenchymal stromal cells (OMSCs).
Exclusion Criteria
-
Review articles, editorials, letters, news articles, and case reports.
-
Studies without a direct focus on MSCs in CNS demyelination.
-
Human clinical trials.
-
Studies lacking experimental models or sufficient data.
Full-text articles were excluded if they provided insufficient data on MSCs or their derivatives in remyelination, lacked experimental evidence, or focused on unrelated topics.
Study Selection Process
The study selection process followed PRISMA guidelines and is summarized in the PRISMA flow diagram (Figure 1). Screening was conducted in three stages: title review, abstract evaluation, and full-text assessment. Titles were initially screened for relevance, followed by abstract evaluation against inclusion criteria. Full-text articles were then assessed for eligibility, with particular attention to the source of cells, study design, treatment methods, experimental models, and reported outcomes. Two independent reviewers conducted all stages of the selection process, and discrepancies were resolved through discussion until consensus was reached (Figure 2).
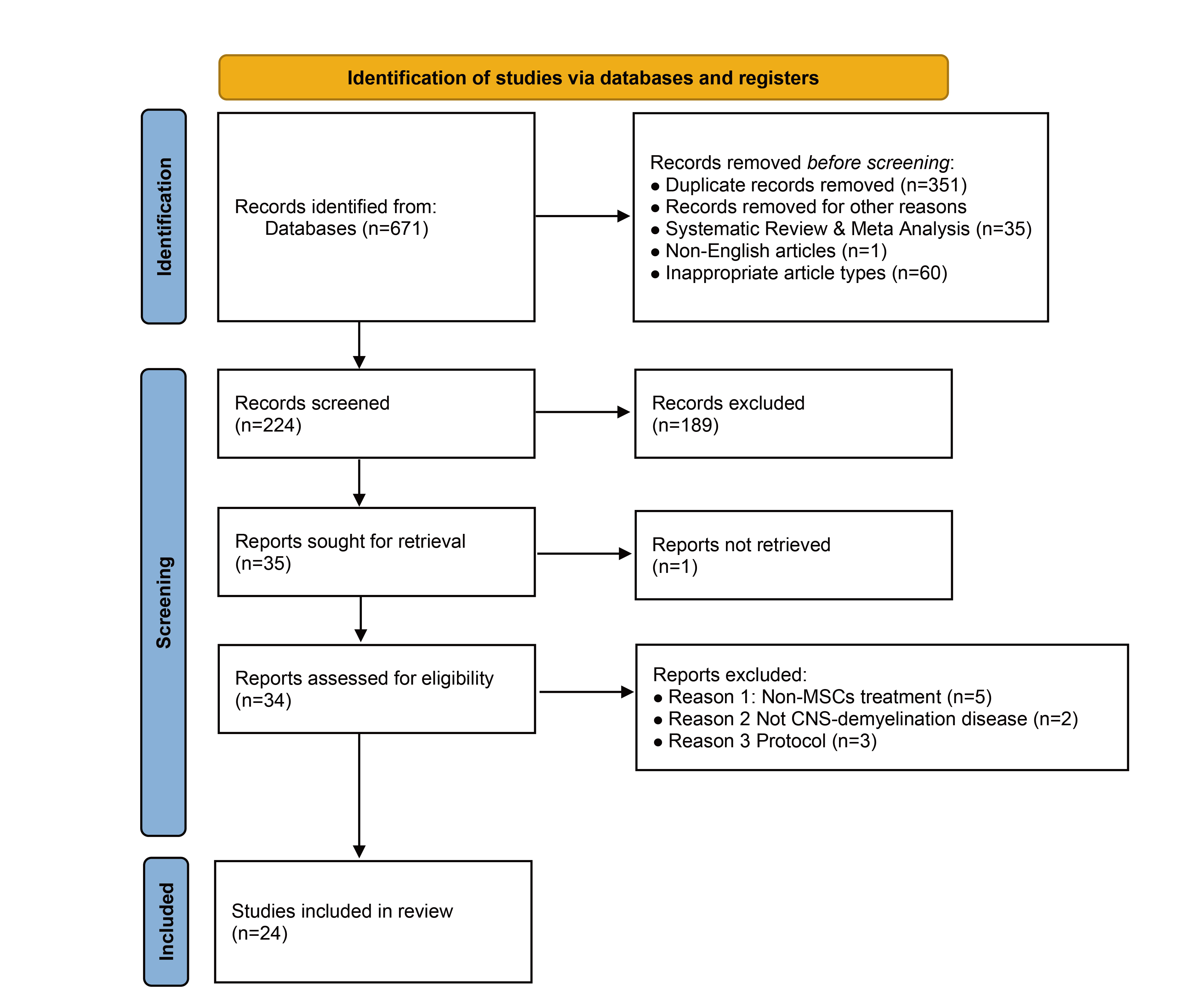
PRISMA flow diagram of the literature selection process. Flowchart detailing the systematic identification, screening, eligibility assessment, and final inclusion of studies. From an initial 671 records, 24 studies were ultimately selected for the review after removing duplicates, ineligible publication types, and records that did not meet the predefined inclusion criteria during full-text assessment.
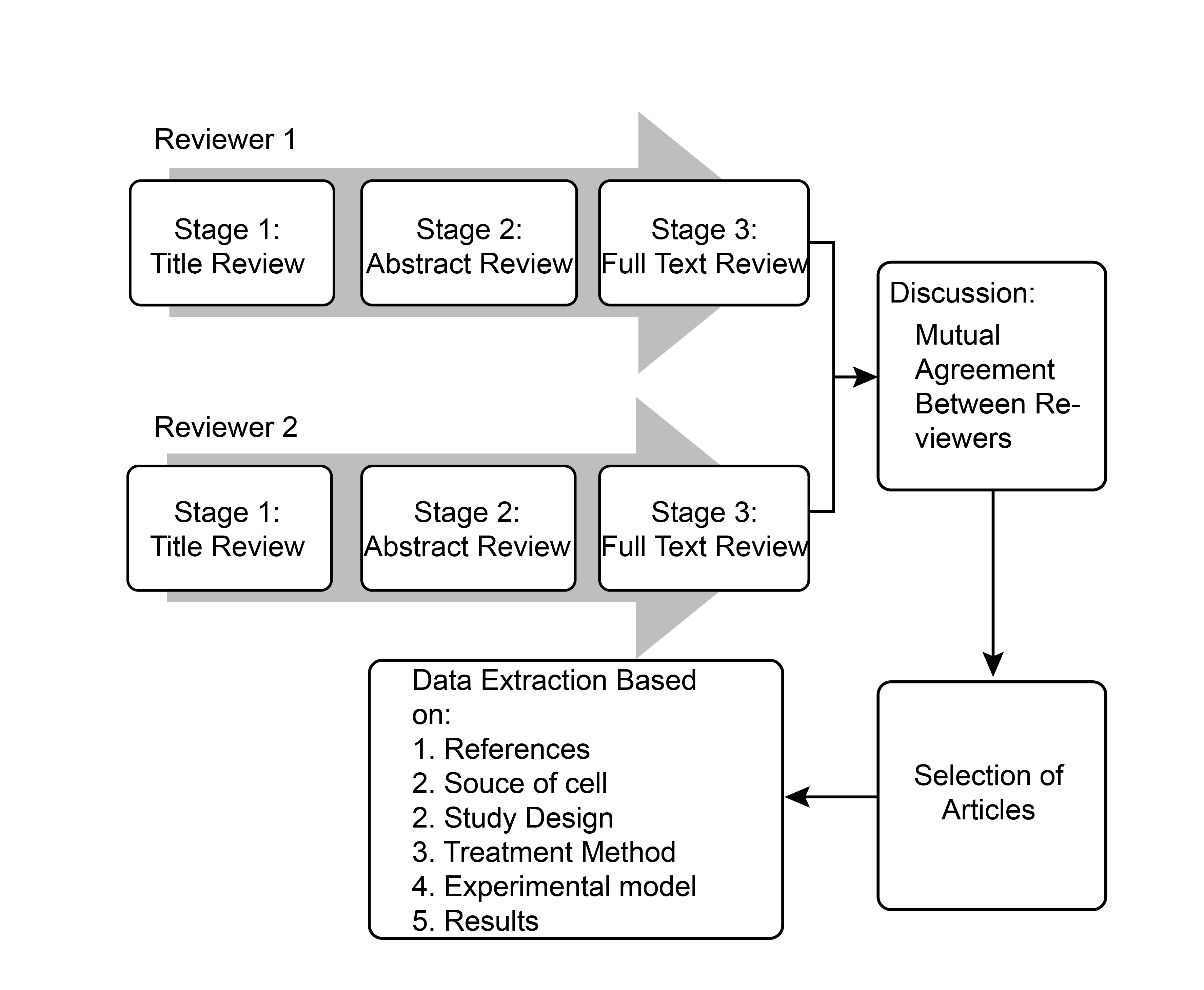
Flow chart of the study selection process. The diagram illustrates the parallel, three-stage screening process conducted independently by two reviewers. The screening involved a title review (Stage 1), followed by an abstract review (Stage 2), and a full-text review (Stage 3). Discrepancies between the reviewers at any stage were resolved through discussion to reach a mutual agreement before the final selection of articles. Following article selection, data extraction was performed based on predefined categories, including references, source of cells, study design, treatment method, experimental model, and results.
Data Extraction
Data were independently extracted by two reviewers using a standardized data collection form (provided as supplementary material). Extracted variables included:
-
Author(s) and year of publication
-
Type of MSC studied
-
Treatment method
-
Study Design (In Vivo or In Vitro)
-
Experimental model
-
Treatment duration
-
Summary of treatment administration, methods, and assessments
-
Markers for oligodendrocyte lineage
-
Main outcomes related to remyelination or OPC differentiation into mature oligodendrocytes (OLs)
All extracted data were compiled and synthesized to identify key findings, mechanistic insights, and gaps in the literature.
Risk of Bias Assessment
Risk of bias in the included studies was evaluated using the Cochrane Risk of Bias Tool, adapted for nonhuman experimental studies. Two independent reviewers assessed domains such as random sequence generation, baseline comparability, allocation concealment, random housing, blinding of caregivers and outcome assessors, and completeness of outcome data. Each domain was rated as low risk, high risk, or unclear risk. Disagreements were resolved through discussion until consensus was reached. (Figure 3, 4)
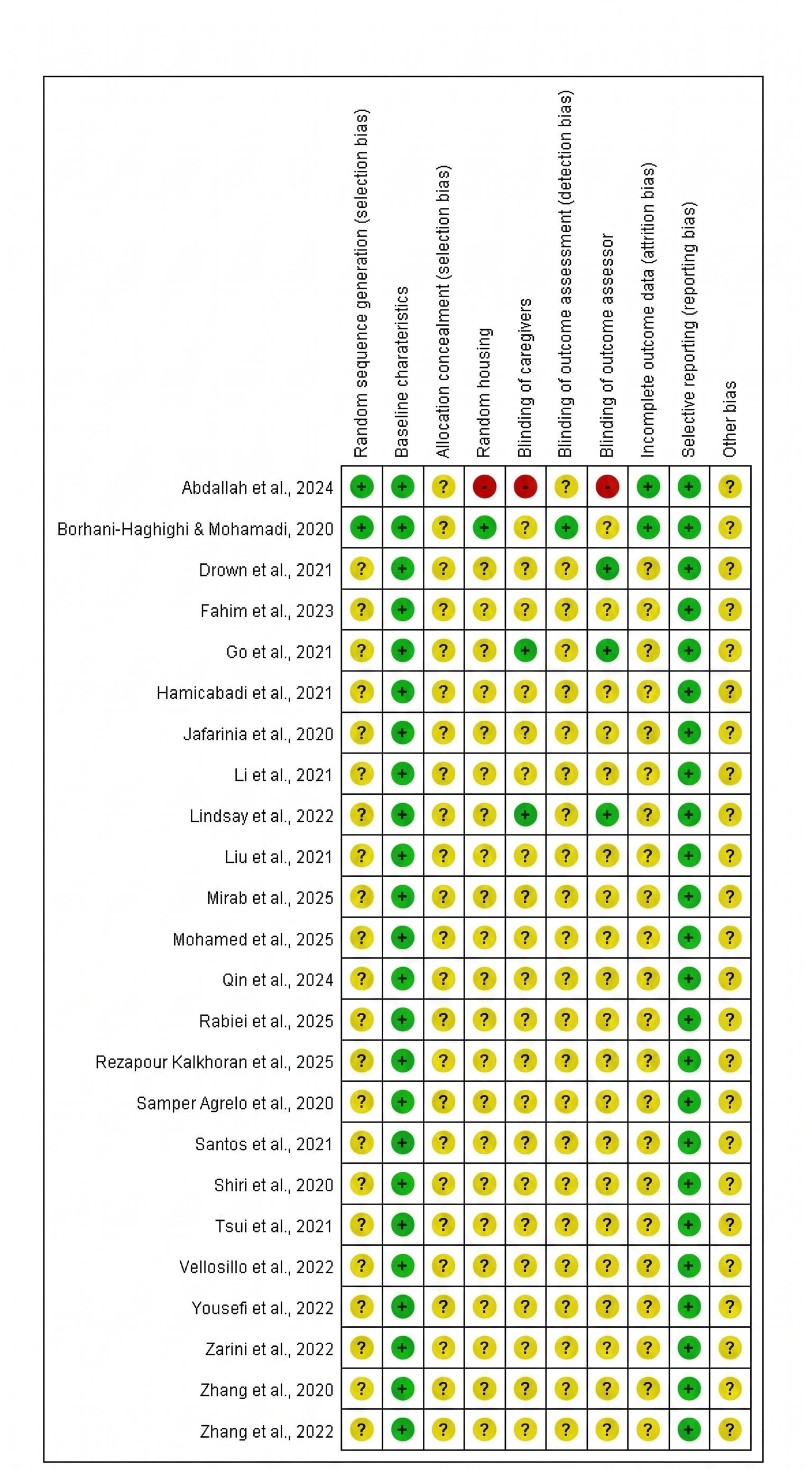
Summary of bias risk for the included studies. The risk of bias was evaluated for each individual study using the Cochrane Risk of Bias Tool adapted for nonhuman experimental studies. The grid presents the independent reviewers' judgments across specific methodological domains, including random sequence generation, baseline characteristics, allocation concealment, random housing, blinding of caregivers, blinding of outcome assessment, blinding of outcome assessor, incomplete outcome data, selective reporting, and other biases. Ratings are categorized as low risk (green '+'), unclear risk (yellow '?'), or high risk (red '-').
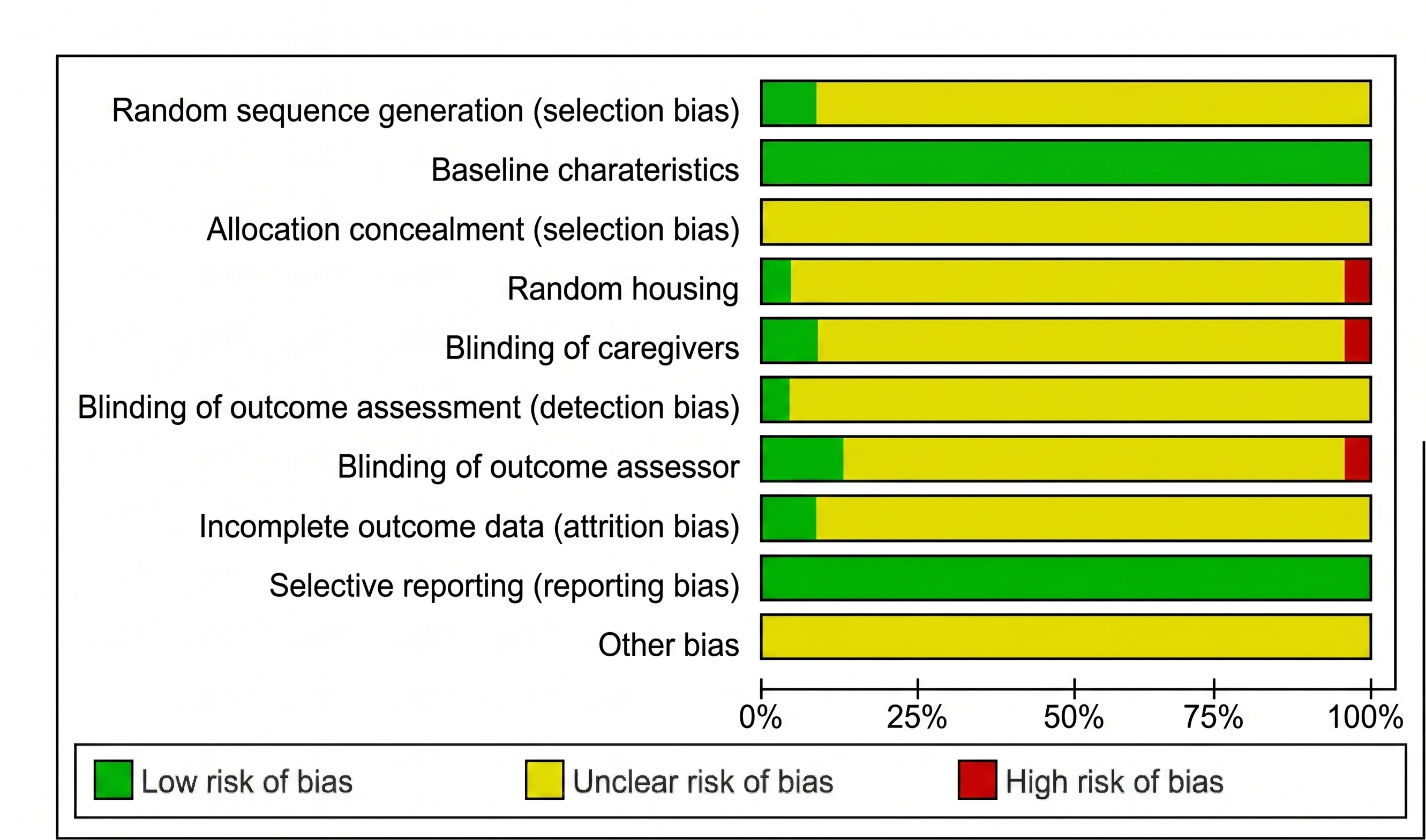
Bias risk graph. This stacked bar chart presents the aggregated risk of bias judgments across all included studies, expressed as percentages. Methodological quality was evaluated using the Cochrane Risk of Bias Tool adapted for nonhuman experimental studies. Each horizontal bar represents a specific methodological domain, illustrating the proportion of studies categorized as having a low risk (green), unclear risk (yellow), or high risk (red) of bias. The graph highlights a high prevalence of "unclear" risk across most domains (such as allocation concealment, random housing, and blinding procedures), primarily due to insufficient methodological reporting in the selected articles, whereas baseline characteristics and selective reporting largely demonstrated a low risk of bias.
Results
Study Selection
The initial database search identified 671 records (PubMed: 200; Scopus: 207; Google Scholar: 264). After removal of 351 duplicates and exclusion of 96 articles (including 35 systematic reviews/meta-analyses, 1 non-English article, and 60 inappropriate article types), 224 unique records remained. Title and abstract screening excluded 189 records that did not meet inclusion criteria. Full-text review was conducted for 34 articles, of which 10 were excluded (5 for non-MSC interventions, 2 for not addressing CNS demyelination, and 3 for being protocols). Ultimately, 24 studies were included in this systematic review (Figure 1).
Characteristics of Included Studies
The included studies investigated MSCs and their derivatives, such as conditioned media (CM), extracellular vesicles (EVs), and MSC-derived oligodendrocyte progenitor cells (OPCs), in remyelination. MSC sources included umbilical cord (UC-MSCs), bone marrow (BM-MSCs), adipose tissue (AD-MSCs), dental pulp (DPSCs), and olfactory mesenchymal stromal cells (OMSCs). Studies involving differentiated MSC-derived cells or secreted products were included when relevant to remyelination mechanisms.
Study Designs and Experimental Models
Of the 24 studies, 21 were in vivo and 3 were in vitro (Figure 3). In vivo studies predominantly used mouse models (n=18), followed by rats (n=2), dogs (n=1), and monkeys (n=1). No human clinical trials were included. (Figure 5) The most frequently studied MSC types in vivo were BM-MSCs (n=9), AD-MSCs (n=6), and UC-MSCs (n=5). Single studies investigated OM-MSCs and DPSCs. In vitro studies similarly focused on AD-MSCs, BM-MSCs, and UC-MSCs, with occasional inclusion of OM-MSCs and DPSCs.
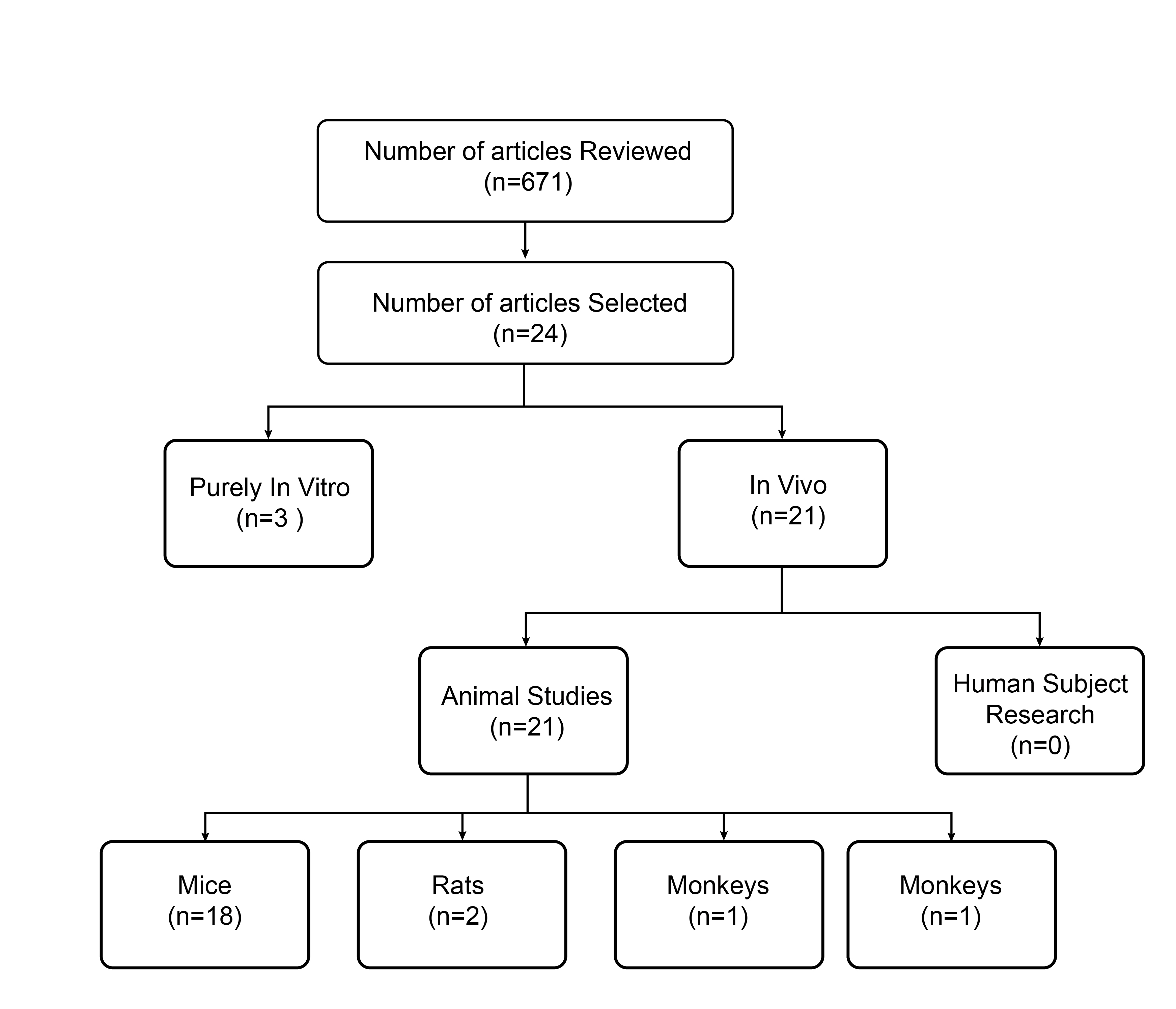
Article distribution by study design and experimental model. The flowchart illustrates the categorization of the 24 selected articles based on their experimental approach. The included studies were broadly divided into purely
Summary of Study Outcomes
Despite heterogeneity in design and treatment protocols, most studies reported that MSCs and their derivatives enhanced remyelination. Key mechanisms included stimulation of OPC proliferation, promotion of OPC differentiation into mature oligodendrocytes (OLs), and modulation of microglial activity. Table 1 summarizes the detailed characteristics and findings of the included studies.
Characteristics of manuscripts reviewed
| Authors. (Year) | Types of MSC | Treatment Method | Study Design | Experimental model | Treatment duration | Summary of Treatment Administration, Methods, and Assessments | Markers of OL lineage | Main Outcomes |
|---|---|---|---|---|---|---|---|---|
| Vellosillo et al. (2022) | rADSC | Induced-OPCs | In vitro | DRG neurons | 5 months |
hADSCs transduced with tetracycline-inducible OL lineage transcription factors (Sox10, Olig2, Zfp536, Nkx6.1). Differentiation assessed by immunocytochemistry. Co-culture with DRG neurons; myelination confirmed via ICC and TEM. |
O4 MAG |
rADSCs effectively converted into OPCs via Sox10, Olig2, Zfp536 expression. |
|
Li et al. (2021) | rBMSC | CM |
In vitro In vivo |
2,5-hexanedion-induced OPCs 2,5-hexanedion-induced demyelinating neurotoxicity model of rats | 5 weeks |
BMSCs administered via tail vein injection. OPCs incubated with HD ± BMSC-CM. Differentiation assessed by immunostaining/ morphometry. Hes1, Notch1, TNFα/RelB measured by ICC and WB. |
NG2 MBP |
BMSC-CM promoted OPC differentiation into mature OLs (in vitro and in vivo). Effects mediated via TNFα/RelB-Hes1 pathway. |
|
Go et al. (2021) | mkBMSC | EV | In vivo | A cortical injury targeting the hand representation area of the primary motor cortex (M1) in a monkey | 14 days |
IV administration of BMSC-EVs. Oligodendrocyte maturation and myelin marker changes in sublesional white matter assessed by IHC, confocal microscopy, stereology, qRT-PCR, ELISA. |
CC1 BCAS1 |
EV treatment increased density of mature CC1+ cells and newly formed myelinating oligodendrocytes (BCAS1++/CC1−, BCAS1++/CC1+). BMSC-EVs reduced myelin damage and promoted myelin maintenance after cortical injury in aged brains. |
|
Qin et al. (2024) | hUCMSC | Exosome |
In vitro In vivo |
OPCs were treated with hUCMSCs-Exos EAE mice | 14 days |
Lentiviral overexpression of miR-23a-3p in hUCMSCs. hUCMSC-derived exosomes used for OPC culture in vitro and administered via tail vein injection in vivo. OPC markers (NG2, PDGFRα) and differentiation markers (MAG, MOG, MBP) assessed by immunofluorescence. Myelin gene expression analyzed by sequencing and bioinformatics. |
NG2 PDGFRα MBP MAG MOG |
hUCMSC-derived exosomes promoted OL-mediated myelinogenesis in MS. Effect mediated via miR-23a-3p transfer, inhibiting Tbr1 activity. PI3K/Akt and Wnt pathways involved in regulation of miR-23a-3p. |
| Fahim et al. (2023) | hUCMSC | Induced OL lineage | In vitro | hUCMSCs overexpressed the transcription factors OLIG2 and MYT1L. | 3 and 7 days |
hUMSCs lineage specification/differentiation assessed by qPCR. Differentiation confirmed via ICC for oligodendrocyte-specific proteins. |
SOX10 NKX2.2 GALC CNP CSPG4 MBP PLP1 |
OLIG2 and MYT1L induced hUMSC differentiation into oligodendrocyte-like cells (gene and protein levels, normal and induction media). |
| Yousefi et al. (2022) | hADSC | hADSCs-overexpressed LIF/IFN‑β |
In vitro In vivo |
hADSCs overexpressing LIF and IFN-β EAE mice | 20 days |
hADSCs engineered to overexpress IFN-β and LIF. Tail vein injection of IFN-β/LIF-overexpressing hADSCs in EAE mice. Effects on CNS immunoregulation and demyelination assessed. |
Olig2 MBP |
Reduced IL-17, IFN-γ, TNF-α; increased IL-10 and IL-4. Increased Olig2+ cell numbers and MBP protein expression Reduced demyelination. |
| Tsui et al. (2021) | rBMSC | Induced OL lineage |
In vitro In vivo |
Chemically induced OPCs from DMSCs Shiever mice | 21 days |
OPCs derived from rat BMSCs using two-step protocol. OPCs co-cultured with neurons in vitro. Induced OPCs injected into mouse corpus callosum in vivo. |
Olig2 NG2 PDGFRα Sox10 |
BMSCs differentiated into OPCs expressing Olig2, NG2, PDGFRα, and Sox10 within 21 days (two-step protocol). OPCs matured into OLs forming compact myelin in vitro and in vivo, without genetic/epigenetic manipulation or viral vectors. |
| Lindsay et al. (2022) | hOMMSC | hOMMSCs |
In vitro In vivo |
Co-culture OM-MSCs with OPCs. EAE mice | 21 days |
Rat microglia and OPCs cultured; OM-MSCs co-cultured with OPCs. hOMMSCs administered via tail vein injection. Comparative efficacy of hOMSCs vs. hBM-MSCs assessed in EAE model. |
O4 PLP |
IL-16 reduced mature PLP+ oligodendrocytes and de novo myelination (in vitro). hOMMSCs promoted therapeutic effects in MS via IL-16-mediated pathway during active demyelination/inflammation (in vivo). OMMSCs more effectively ameliorated severe EAE progression compared to BMSCs. |
| Zhang et al. (2020) | rBMSC | rBMSCs | In vivo | EAE mice | 28days |
BM-MSCs (P2–P3) injected into ventricles. OL lineage markers (A2B5, O4, CNPase) detected by IF staining at multiple time points post-injection. |
A2B5 O4 CNPase |
Increased A2B5+, O4+, and CNPase+ cell numbers from day 3, sustained through days 7–28 post-treatment. MSC treatment promoted oligodendrocyte lineage cell expression. |
| Samper Agrelo et al. (2020) | rBMSC | CM |
In vitro In vivo |
aNSCs Female rats and mice | 28days |
aNSCs pre-stimulated with MSC-CM. Injected into corpus callosum and cortex of mouse brain; transplanted into rat spinal cord grey/white matter. Secretome analyzed by mass spectrometry. |
NG2 Olig2 GSTπ MBP |
Increased NG2, Olig2, GSTπ, MBP expression; decreased GFAP+ cells. TIMP-1 in MSC-CM promoted oligodendrocyte differentiation and myelination. MSC-secreted factors enhanced oligodendrogenesis from primary aNSCs and OPCs. |
| Zhang et al. (2022) | mkBMSC | Exosome | In vivo |
EAE mice CPZ mice |
4 weeks for EAE 2 weeks for CPZ |
BM-MSC-derived exosomes administered via tail vein injection. MBP protein levels measured by WB. OPC proliferation (BrdU-NG2) and differentiation (BrdU-APC) assessed by double IF staining. Microglial function and neuroinflammation evaluated by IF staining and WB. |
MBP NG2 APC |
BMSC-Exo improved neurological outcomes. Increased newly generated oligodendrocytes (BrdU+/APC+), mature oligodendrocytes (APC+), and MBP levels. Reduced APP+ density. Alleviated neuroinflammation by promoting M2 and suppressing M1 microglial phenotypes, with cytokine modulation. Inhibited TLR2/IRAK1/NFκB pathway. Promoted remyelination and reduced neuroinflammation in CNS. |
| Shiri et al. (2021) | hUCMSC | hUCMSCs | In vivo | CPZ mice | 2weeks |
UCMSCs injected into lateral ventricle. Demyelination/remyelination assessed in corpus callosum by LFB staining. Oxidative stress markers (MDA, GSH, SOD) measured post-cuprizone induction. Mitochondrial/metabolic transcription factors (PGC1α, NRF-1, TFAM, MFN2, DRP1) quantified. | Olig2 |
MSC transplantation increased oligodendrocyte numbers and reduced astrogliosis/microgliosis. MSCs alleviated MS by promoting antioxidant response, reducing oxidative stress, and improving mitochondrial homeostasis. |
| Brown et al. (2021) | hUCMSC | OL lineage |
In vitro In vivo |
NSCs EAE mice | 2 weeks |
MSCs differentiated into NSCs; transplanted via tail vein injection. Immune infiltrates assessed by HE staining and IHC; Treg/Th17 levels analyzed by flow cytometry. Myelination evaluated by LFB staining and immunostaining. Transplanted cell fate and expression of inflammation, astrogliosis, myelination, neural differentiation, neuroprotection, and neurogenesis markers investigated by IHC and qRT-PCR. |
NESTIN TUJ1 VIMENTIN PAX6 OLIG2 SOX10 O4 MBP MOG |
UCMSC-derived NSCs expressed neural markers (NESTIN, TUJ1, VIMENTIN, PAX6). UCMSCs differentiated into NSCs and oligodendrocytes, expressing OLIG2, SOX10, O4, MBP, MOG. NSCs modulated Treg/Th17 balance. Upregulated myelination/neuroprotection genes; downregulated inflammation/astrogliosis genes. Induced anti-inflammatory response; promoted myelination, neuroprotection, and neurogenesis in EAE models. |
| Santos et al. (2022) | hADSC | OL lineage | In vitro | Neuron-like cells and oligodendrocyte-like cells derived from hADSCs were co-cultured | 7 days |
hADSCs differentiated into NSCs; co-cultured with oligodendrocyte-like and neuron-like cells. Proliferation assessed by BrdU incorporation. In vitro myelination evaluated by gene expression (Mbp, Sox10, Map2) and IF labeling (O4/GalC, MAP2). |
Nestin GalC O4 |
NSCs expressed nestin; astrocytes GFAP; oligodendrocytes GalC/O4 (immunostaining). Co-cultured NLCs/OLCs showed excitability (81.3%); 23.5% expressed overlapping neuron/oligodendrocyte markers, indicating in vitro myelination. hADSCs transdifferentiated into NSCs and neural phenotypes, with in vitro neuron myelination observed. |
| Liu et al. (2021) | mBMSC | OL lineage |
In vitro In vivo |
Differentiation of BMSCs EAE mice | 28 days |
BMSCs injected into lateral ventricle. Differentiation in hippocampus/cortex assessed by IF. Disease incidence, neurobehavioral scores, and weight monitored in EAE and BMSC-transplanted mice. Inflammatory infiltration and demyelination evaluated by HE and LFB staining. Serum inflammatory factors measured by ELISA; spinal cord mRNA expression analyzed by RT-qPCR. | MBP |
BMSCs differentiated into neurons, astrocytes, and oligodendrocytes (in vitro and in vivo). Transplantation reduced TNF-α, IL-1β, IFN-γ; increased IL-10 and TGF-β. Alleviated inflammation and demyelination in EAE mice, potentially via neuronal, astrocytic, and oligodendrocytic differentiation. |
| Borhani-Haghighi & Mohamadi. (2020) | hUCMSC | CM |
In vitro In vivo |
Conditioned medium was harvested from the induced OLs EAE mice | 28 days |
UCMSCs induced to differentiate into OLs; identity confirmed by ICC. OL-conditioned medium harvested after 72 h culture and administered intranasally to EAE mice. Lumbar spinal cords collected at day 28; analyzed by WB, IHC, RT-PCR, and histochemistry. |
MBP Olig2 O4 |
OL-CM contained BDNF, GDNF, CNTF; reduced inflammation, promoted remyelination, and restored body weight. Improved neurological function in EAE mice via anti-inflammatory and regenerative effects. Intranasal OL-CM (from WJMSC-derived OLs) proposed as novel MS therapy. |
| Hamidabadi et al. (2021) | hDPSC | OL lineage |
In vitro In vivo |
Differentiation of hDPSCs LPC-induced demyelination mouse model | 2 weeks |
LPC stereotaxic injection into corpus callosum to establish demyelination model in female Sprague-Dawley rats. hDPSCs transplanted into corpus callosum via stereotaxic injection. Maturation of transplanted hDPSCs evaluated. |
Olig2 O4 PLP |
Differentiated OPCs confirmed by Olig2 and O4 immunofluorescence. Engraftment decreased demyelination and increased remyelination. PLP immunohistochemistry confirmed mature myelinating oligodendrocytes. hDPSCs transdifferentiated into oligoprogenitor cells in vivo. |
| Jafarinia et al. (2020) | hADSC | EV | In vivo | EAE mice | 20 days |
hADSC-EVs administered intravenously. Leukocyte infiltration, demyelination, and MFI of MBP/OLIG2 evaluated by histology and IHC. |
MBP OLIG2 |
hADSC/hADSC-EV treatment reduced clinical score and MOG-induced splenocyte proliferation in mice. hADSC-EVs attenuated EAE by reducing T-cell proliferation, clinical score, leukocyte infiltration, and demyelination. |
| Zarini et al. (2022) | mBMSC | BMSC | In vivo | CPZ mice | 2 weeks |
MSCs administered intranasally. Outcomes evaluated by LFB staining, Rotarod test, qRT-PCR, IHC, and WB. | APC |
MSCs improved motor coordination and promoted myelin recovery (intermittent cuprizone model). Inhibited astrogliosis/microgliosis; increased mature oligodendrocytes. Reduced M1 macrophage markers; promoted M2 macrophage recovery. Decreased pro-inflammatory markers; increased anti-inflammatory markers. Intranasal MSC delivery reshaped macrophage polarity and modulated glial, inflammatory, and angiogenic markers, supporting therapeutic outcomes. |
| Abdallah et al. (2024) | dADSC | dADSC and exosomes | In vivo | Ethidium bromide dogs | 3, 7, 14, and 28 days |
Stem cells or exosomes injected intrathecally via foramen magnum into proximal spinal cistern (dog MS model). Single intrathecal treatment administered. Outcomes evaluated clinically, radiologically, histopathologically, by electron microscopy, and CSF analysis. |
Treated groups showed improved clinical signs vs. controls. MRI confirmed resolution of hyperintense lesions. Histopathology/electron microscopy revealed healing and remyelination; stem cells more effective than exosomes. | |
| Rezapour Kalkhoran et al. (2025) | hADSC | MSCs | In vivo | EAE mice | 23 days |
MSCs (or treatment) administered intravenously via tail vein in EAE mice. Inflammation and demyelination assessed by HE and LFB staining. Oligodendrocyte lineage and maturation markers (Olig2, Nogo-A) detected by IHC in brain and spinal cord. |
Olig2 Nogo-A |
s-KL-MSCs alleviated EAE clinical signs more effectively than unmodified MSCs. Reduced CNS inflammation and demyelination. Increased Olig2 and Nogo-A mRNA/protein expression, enhancing oligodendrocyte differentiation and maturation. |
| Rabiei et al. (2025) | rBMSC | Exosome | In vivo | CPZ mice | 2 weeks |
Behavioral outcomes assessed by Y-maze, novel arm discrimination, and wire hang tests. Myelin integrity evaluated by FluoroMyelin, Black-Gold II staining, and MOG immunofluorescence. Gene expression of myelin (MBP, Olig2, PDGFRα), antioxidant (Nrf2, HO-1, Nqo1), and inflammatory (IL-1β, IL-18, TNF-α) markers analyzed by qRT-PCR. Oxidative stress (MDA, TAC, SOD, CAT, GPx) and glial activation (Iba1, GFAP) markers assessed. |
Olig2 MBP PDGFRα |
EXO+KMP therapy improved working memory, spatial recognition, and motor endurance in CPZ mice. Enhanced myelin integrity and reduced demyelinated regions (corpus callosum). Upregulated MBP and Olig2; downregulated OPC marker PDGFRα. Activated Nrf2/HO-1/Nqo1 pathway; increased TAC and antioxidant enzymes; reduced MDA. Decreased microglial (Iba1) and astrocytic (GFAP) activation; suppressed IL-1β, IL-18, TNF-α. |
| Mirab et al. (2025) | hUCMSC | Exosome | In vivo | CPZ mice | 3 weeks |
Motor coordination assessed weekly by beam walking test. Brain tissues collected post-sacrifice; analyzed by LFB/PAS staining, IHC, and qRT-PCR. IHC for MBP, Iba-1, GFAP; qRT-PCR for oligodendrocyte lineage genes (Olig2, Sox10, Plp, MBP). |
Olig2 Sox10 Plp MBP |
hUCMSC-Exos improved motor performance in CPZ mice (BWT). Increased myelin repair in corpus callosum (histology). Reduced demyelination (immunohistochemistry). Decreased microglial and astrocytic activation. Upregulated genes linked to oligodendrocyte differentiation and maturation. |
| Mohamed et al. (2025) | rADSC | rADSC | In vivo | CPZ mice | 6 weeks |
MSCs (or treatment) administered intranasally Motor function assessed by wire hang test at week 6. Outcomes evaluated by HE staining, IHC (Olig2, Iba-1), TEM, and histomorphometric analysis. | Olig2 |
ADMSCs ameliorated histological alterations in corpus callosum after CPZ-induced demyelination. Increased OPCs via enhanced Olig2 expression. Reduced active microglia by decreasing Iba-1 expression. |
Discussion
The Potential of Mesenchymal Stem Cells for Multiple Sclerosis Therapy
Wharton’s jelly-derived MSCs (WJ-MSCs) and adipose-derived MSCs (AD-MSCs) demonstrate the most promising profiles for MS therapy 22,23. WJ-MSCs are advantageous due to their non-invasive and ethically acceptable collection, high proliferative capacity, potent immunomodulatory effects (e.g., inhibition of Th1/Th17 responses and promotion of Tregs), low immunogenicity, and proven safety in large-scale allogeneic trials 24. AD-MSCs similarly offer abundant availability, high expansion rates, and strong immunosuppressive properties 25. In contrast, bone marrow-derived MSCs (BM-MSCs) face challenges such as invasive harvesting, low yield, and age-related decline in potency. Other MSC sources (umbilical cord blood, dental pulp, olfactory mucosa, placenta, menstrual blood) provide unique benefits but often lack scalability or consistency 26,27.
Developmental Stages and Marker Dynamics of the Oligodendrocyte Lineage
Oligodendrocyte development follows a defined lineage, beginning with neural progenitor cells (NPCs), which differentiate into oligodendrocyte progenitor cells (OPCs) and subsequently into mature oligodendrocytes (OLs) 28. NPCs are identified by markers such as Nestin and Sox2 29,30, while OPCs express transcription factors (Olig2, Sox10, Nkx2.2) and surface antigens (A2B5, NG2, PDGFRα, O4) 31,32,33. Mature OLs are characterized by markers including GalC, MBP, MAG, and CC1 34,35. Combinations of markers, such as BCAS1 and CC1, improve specificity in distinguishing maturation stages 36. Understanding these marker dynamics is critical for evaluating remyelination and optimizing MSC-based therapies (Figure 6)
MSCs and Direct Remyel ination
Seven studies (five in vivo and two combining in vitro and in vivo approaches) demonstrated that MSCs directly promote remyelination in experimental autoimmune encephalomyelitis (EAE) and cuprizone (CPZ) models 37,38,39,40,41,42,43,44. Delivery methods included tail vein injection, ventricular injection, and intranasal administration, each with distinct advantages and limitations. Tail vein injection enables systemic distribution but is hindered by pulmonary entrapment, while ventricular injection provides direct access to cerebrospinal fluid but is invasive. Intranasal administration is non-invasive and bypasses the blood-brain barrier but has variable efficiency. A combined approach, such as ventricular priming followed by tail vein maintenance, may optimize efficacy and safety.
MSCs enhanced OPC proliferation, migration, and differentiation into mature OLs via multiple mechanisms. For example, AD-MSCs and UC-MSCs increased Olig2+ and MBP+ cells, while AD-MSCs overexpressing INF-β and LIF modulated inflammation 38,41. BM-MSCs promoted lineage expansion and reduced astrogliosis and microgliosis 40, while olfactory MSCs demonstrated superior efficacy in severe EAE models via IL-16-mediated pathways 39. However, standardized protocols and direct comparisons of delivery methods remain lacking, and many studies rely on single-marker quantification.
Effects of MSC-Derived Conditioned Medium and Extracellular Vesicles
The paracrine activity of MSCs, mediated through conditioned medium (CM) and extracellular vesicles (EVs), offers promising cell-free alternatives to whole-cell therapies. CM contains soluble factors such as cytokines and growth factors, while EVs deliver bioactive molecules including proteins, lipids, and nucleic acids 45,46. Both approaches have shown efficacy in demyelination models, enhancing OPC differentiation, promoting myelin regeneration, and modulating inflammatory pathways.41,42,43,44,45,46,47,48,49.
For example, BM-MSC-CM promoted OPC differentiation via the TNFα/RelB-Hes1 pathway 48,50. while UC-MSC-derived exosomes overexpressing miR-23a-3p activated the PI3K/Akt pathway and inhibited Wnt5a to promote remyelination 49. In cuprizone models, UC-MSC exosomes improved motor function, enhanced corpus callosum remyelination, and reduced gliosis 52. Synergistic effects were observed when MSC-derived exosomes were combined with kaempferol, yielding improved motor and cognitive outcomes and reduced oxidative stress 53. Despite these findings, long-term efficacy, scalability, and direct comparisons with whole-cell MSC therapies remain unclear.
MSC-Derived Oligodendrocyte Lineage Cells
MSC differentiation into oligodendrocyte lineage cells offers another therapeutic avenue. Seven studies explored MSC-derived OPCs and OLs using transcription factor overexpression (e.g., Sox10, Olig2, Zfp536) and chemical induction (e.g., SHH, T3) 52,53,54,55,56. AD-MSCs converted via Sox10, Olig2, and Zfp536 expressed mature OL markers and successfully ensheathed DRG axons 61, while UC-MSCs differentiated with OLIG2/MYT1L or SHH/T3 yielded OLs expressing MBP and MOG, alongside immunomodulatory effects 59,62. Additionally, rat BM-MSCs and human DPSCs demonstrated the ability to generate myelinating OLs both in vitro and in vivo 56,58,60.
Despite these advances, key challenges remain. Differentiation efficiency is generally high but varies depending on MSC source and protocol. AD-MSC-based protocols achieve over 95% A2B5+ and Olig2+ cells, with approximately 20% MBP+ OLs and minimal off-target gliogenesis (GFAP+ <1%) 19. UC-MSCs yield nearly all marker-positive cells (A2B5, O4, GalC, MBP) with robust proliferation 63, while TET3-mediated UC-MSC differentiation rapidly upregulates OPC markers (NG2, PDGFRA) within 7 days 64. However, long-term stability and the efficiency of aged MSCs remain underexplored.
Clinical translation faces significant hurdles. Differentiation success, purity, and stability require further validation. Key risks include tumorigenesis, reduced cell survival in inflammatory environments, and the potential for viral mutagenesis. Prolonged MSC culture may lead to senescence or transformation into cancer-associated fibroblasts, while exposure to IFN-γ can impair cell viability and increase immunogenicity. Although lentiviral vectors are effective, they carry the risk of insertional mutations 65. Furthermore, direct comparisons with whole MSCs or exosomes are lacking, which may represent complementary or safer therapeutic alternatives. Future studies should prioritize optimizing differentiation protocols, mitigating risks, and evaluating therapeutic efficacy in demyelination models.
Functional Recovery and Translational Relevance
While MSC-based therapies consistently demonstrate remyelination at histological and molecular levels, functional recovery outcomes remain underexplored. Some studies reported improved motor coordination and cognitive performance in CPZ models using behavioural tests such as beam walking, Y-maze, and Rotarod 52,53,66. However, many studies focus primarily on cellular markers without assessing functional outcomes 39,47, highlighting a critical gap. Future research should incorporate standardized functional assessments to better evaluate translational potential.
Heterogeneity and Publication Bias
Considerable heterogeneity exists across studies in terms of MSC sources, experimental models, delivery methods, and outcome measures. This variability complicates direct comparisons and limits the generalizability of findings. Moreover, the predominance of positive results raises concerns about publication bias, which may lead to an overestimation of MSC efficacy. Greater transparency, including the reporting of negative or inconclusive findings, is essential to provide a balanced understanding of MSC-based therapies.
Methodological Limitations and Risk of Bias
Several studies lacked detailed reporting of key methodological practices such as randomization, allocation concealment, and blinding of outcome assessors. These omissions hinder comprehensive risk-of-bias assessment and reduce confidence in the reproducibility of results. Future research should adhere to established guidelines, such as ARRIVE and SYRCLE, to improve methodological rigor and ensure more reliable preclinical evidence.
Critical Appraisal of Evidence Consistency and Strength
The evidence synthesized in this review highlights the potential of MSCs to promote remyelination through direct transplantation, paracrine activity via conditioned medium (CM) and extracellular vesicles (EVs), and differentiation into oligodendrocyte lineage cells. However, the consistency and strength of this evidence vary considerably. For instance, while direct MSC transplantation has demonstrated benefits in OPC proliferation and differentiation, delivery methods such as tail vein, ventricular, and intranasal administration have not been directly compared, and variability in MSC types, doses, and experimental conditions complicates interpretation.
Similarly, MSC-derived CM and EVs show promise as cell-free alternatives, but their relative efficacy compared to whole-cell therapies remains unclear, and long-term stability and scalability are underexplored. Differentiation of MSCs into oligodendrocyte lineage cells represents a promising avenue for targeted remyelination, yet this approach requires complex protocols and further validation to ensure safety, functional integration, and durability. Challenges such as immune responses, senescence, and tumorigenicity must be addressed before clinical translation can be realized.
Study Limitations
This review has several limitations. First, significant heterogeneity exists across the included studies, encompassing diverse MSC sources, derivatives, animal models, and outcome measures. This variability limits generalizability and complicates direct comparisons. Second, the absence of human clinical trials and the limited evaluation of functional recovery outcomes such as motor, sensory, and cognitive improvements which restrict the clinical applicability of findings. Third, the predominance of positive results raises concerns about publication bias, potentially leading to an overestimation of MSC efficacy. Finally, many studies lacked transparency in reporting methodological details such as randomization and blinding, hindering comprehensive risk-of-bias assessment and reducing confidence in reproducibility.
Future Directions
Future research should prioritize direct comparisons of MSC sources, derivatives, and delivery methods to identify the most effective strategies. Standardized protocols for MSC and EV isolation, characterization, and therapeutic application are needed to ensure reproducibility. Incorporating functional assessments in preclinical models will bridge the gap between histological and behavioral outcomes. Expansion of well-designed human clinical trials, with diverse patient populations and standardized outcome measures, is essential to enhance clinical relevance. Finally, encouraging the publication of both positive and negative findings will help mitigate publication bias and provide a balanced evidence base.
Conclusion
This systematic review highlights the potential of MSCs and their derivatives including conditioned medium (CM), extracellular vesicles (EVs), and induced oligodendrocyte lineage cells for promoting remyelination in demyelinating diseases. Preclinical studies demonstrate promising results, suggesting their therapeutic potential (Figure 7). However, challenges remain, including heterogeneity of study designs, lack of standardized protocols, and limited human clinical data. Addressing these limitations through rigorous research, methodological standardization, and well-designed clinical trials will be essential for translating MSC-based therapies into safe and effective treatments for multiple sclerosis and related disorders.
Abbreviations
A2B5: Neural surface antigen A2B5; AD-MSCs / ADSCs: Adipose-Derived Mesenchymal Stem Cells; AHSCT: Autologous Hematopoietic Stem Cell Transplantation; AI: Artificial Intelligence; Akt: Protein Kinase B; ARRIVE: Animal Research: Reporting of In Vivo Experiments; BCAS1: Breast Carcinoma Amplified Sequence 1; BM-MSCs / BMSCs: Bone Marrow-Derived Mesenchymal Stem Cells; CC1: Anti-Adenomatous Polyposis Coli (APC) clone CC1; CM: Conditioned Media / Medium; CNS: Central Nervous System; CPZ: Cuprizone; DMTs: Disease-Modifying Therapies; DPSCs: Dental Pulp Stem Cells; DRG: Dorsal Root Ganglion; EAE: Experimental Autoimmune Encephalomyelitis; ESCs: Embryonic Stem Cells; EVs: Extracellular Vesicles; GalC: Galactocerebroside; GFAP: Glial Fibrillary Acidic Protein; Hes1: Hes Family BHLH Transcription Factor 1; IFN-γ: Interferon-gamma; IL-16: Interleukin-16; INF-β: Interferon-beta; iPSCs: Induced Pluripotent Stem Cells; LIF: Leukemia Inhibitory Factor; MAG: Myelin-Associated Glycoprotein; MBP: Myelin Basic Protein; miR-23a-3p: MicroRNA 23a-3p; MOG: Myelin Oligodendrocyte Glycoprotein; MS: Multiple Sclerosis; MSC-CM: Mesenchymal Stem Cell-Conditioned Media; MSCs: Mesenchymal Stem Cells; MYT1L: Myelin Transcription Factor 1-Like; NG2: Neural/Glial Antigen 2; Nkx2.2: NK2 Homeobox 2; NPCs: Neural Progenitor Cells; O4: Oligodendrocyte Marker O4; Olig2 / OLIG2: Oligodendrocyte Transcription Factor 2; OLs: Mature Oligodendrocytes; OM-MSCs / OMSCs: Olfactory Mesenchymal Stromal Cells; OPC / OPCs: Oligodendrocyte Progenitor Cell(s); PDGFRα / PDGFRA: Platelet-Derived Growth Factor Receptor Alpha; PI3K: Phosphoinositide 3-kinase; PRISMA: Preferred Reporting Items for Systematic Reviews and Meta-Analyses; RelB: RELB Proto-Oncogene; SHH: Sonic Hedgehog; Sox2: SRY-Box Transcription Factor 2; Sox10: SRY-Box Transcription Factor 10; SYRCLE: Systematic Review Centre for Laboratory animal Experimentation; T3: Triiodothyronine; TET3: Tet Methylcytosine Dioxygenase 3; Th1 / Th17: Type 1 / Type 17 T helper cells; TNFα: Tumor Necrosis Factor-alpha; Tregs: Regulatory T cells; UC-MSCs / UCMSCs: Umbilical Cord-Derived Mesenchymal Stem Cells; WJ-MSCs: Wharton’s Jelly-Derived Mesenchymal Stem Cells; Wnt5a: Wnt Family Member 5A; Zfp536: Zinc Finger Protein 536
Acknowledgments
None.
Author’s contributions
All authors equally contributed to this work, read and approved the final manuscript.
Funding
None
Availability of data and materials
Not applicable.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Declaration of generative AI and AI-assisted technologies in the writing process
The authors declare that they have not used generative AI (a type of artificial intelligence technology that can produce various types of content including text, imagery, audio and synthetic data).
Competing interests
The authors declare that they have no competing interests.