

In vitro evaluation of the antiproliferative and pro-apoptotic effects of paclitaxel and Illicium verum Hook f. extract in PC3 and LNCaP human prostate cancer cells
- Student Research Committee, Kermanshah University of Medical Sciences, Kermanshah, Iran
- Fertility and Infertility Research Center, Health Technology Institute, Kermanshah University of Medical Sciences, Kermanshah, Iran
- Department of Tissue Engineering, School of Medicine, Kermanshah University of Medical Sciences, Kermanshah, Iran
Abstract
Introduction: Prostate cancer remains one of the most common malignancies among men worldwide. Combining plant-derived compounds with conventional chemotherapeutic agents has attracted increasing attention as a potential strategy to improve anticancer responses. Illicium verum Hook. f. is a traditional medicinal plant with documented biological activities. This study aimed to evaluate the antiproliferative and pro-apoptotic effects of paclitaxel (PTX) and a crude hydroalcoholic I. verum extract (IVE), alone and in combination, in PC3 and LNCaP human prostate cancer cells.
Methods: PC3 and LNCaP cells were treated with IVE at concentrations of 0, 12.5, 25, 50, and 100 µg/mL, and PTX at 0, 5, 10, 25, 50, and 100 nM for 24, 48, and 72 h. Cell proliferation was assessed using the MTT assay, and half-maximal inhibitory concentration (IC50) values were estimated from dose–response curves using GraphPad Prism software. Cells were then treated for 48 h with the IC50 concentrations of PTX, IVE, or their combination, and the mRNA expression levels of P53, BAX, BCL-2, Caspase-3, and Caspase-9 were evaluated using quantitative real-time PCR (qRT-PCR).
Results: Both PTX and IVE significantly reduced the proliferation of PC3 and LNCaP cells in a time- and dose-dependent manner. The estimated 48-hour IC50 values for PTX were 12.5 nM in PC3 cells and 8.1 nM in LNCaP cells, while the corresponding values for IVE were 11.8 µg/mL and 7.1 µg/mL, respectively. Compared with single treatments, the PTX–IVE co-treatment significantly enhanced the expression of the pro-apoptotic P53, BAX, Caspase-3, and Caspase-9 genes while reducing BCL-2 expression.
Conclusion: These findings suggest that IVE may enhance the PTX-associated antiproliferative and pro-apoptotic responses in prostate cancer cells. However, because formal synergy analysis and phytochemical standardization of the extract were not performed, these results should be interpreted as preliminary in vitro screening findings that require further mechanistic and phytochemical validation.
Introduction
Prostate cancer (PC) is one of the most prevalent cancers among men worldwide, ranking as the second most common malignancy after lung cancer and the fifth leading cause of cancer-related mortality in the male population1. According to the GLOBOCAN database report, the mortality rate of PC has been significantly increasing, with 1,466,680 newly diagnosed cases and 396,792 deaths reported in 20221. PC is a complex, multifactorial, and hormonally dependent malignancy that mainly impacts middle-aged men (aged 45–60 years)2. The primary risk factors for PC include age, race, genetic inheritance, germline mutations, sexual activity, obesity, smoking, lifestyle, dietary habits, estrogenic compounds, oxidative stress, and environmental factors3.
Standard diagnostic methods for PC include prostate gland biopsy, measurement of prostate-specific antigen (PSA) levels, immunohistochemistry, transrectal ultrasound, magnetic resonance imaging (MRI), and positron emission tomography (PET) scans4. Current conventional treatments for PC include surgery, chemotherapy, radiotherapy, hormonal therapy, and cryotherapy5. Paclitaxel (PTX) is a widely used chemotherapeutic agent administered for treating several cancers, including lung, breast, ovarian, and prostate cancers. PTX is a well-known antitumor, anticancer, and antimitotic agent with distinct mechanisms of action, approved by the United States FDA (Food and Drug Administration) in 19926. Chemically, PTX is a tricyclic diterpenoid compound (molecular formula: C47H51NO14), initially derived from the bark and needles of Taxus brevifolia (Pacific yew tree) in 19717.
As an antitumor agent, PTX inhibits cell cycle progression, mitosis, and the growth of cancer cells by disrupting tubulin assembly into microtubules, thereby preventing cell division1,2. However, the main drawbacks of PTX in PC treatment are its various side effects and the development of drug resistance, even following short-term therapy3. Chemotherapy drug resistance and extensive adverse effects (including nausea, vomiting, hair loss, headache, dizziness, mucositis, neutropenia, diarrhea, chest pain, leukopenia, anemia, joint pain, hypersensitivity reactions, arthralgia, myalgia, weakness, and neuropathy) lead to therapeutic failure and reduced efficacy4. Consequently, combining chemotherapy strategies with plant-derived herbal compounds (phytochemicals) has recently attracted significant attention in cancer treatment studies.
Phytochemicals are pharmacologically active secondary metabolites extracted from dietary and traditional medicinal plants, possessing potent antioxidant, anticancer, antiproliferative, and anti-inflammatory properties5,6. It has been shown that co-treatment with phytochemicals may enhance chemotherapy-associated anticancer responses and potentially reduce drug resistance or adverse effects7,8. Illicium verum Hook. f. (IV), commonly referred to as star anise or Chinese anise, is an aromatic evergreen tree belonging to the family Magnoliaceae and genus Illicium, with regional distribution in East and Southeast Asia, and southeastern North America9. Star anise fruits contain various phytochemicals, including volatiles, sesquiterpenes, phenylpropanoids, lignans, and flavonoids10.
Due to its rich phytochemical composition, star anise exhibits potent antioxidant, anti-inflammatory, antibacterial, and anticancer properties11. Several studies have investigated the effect of IV extract (IVE) and its bioactive components in suppressing various cancers. Methanolic IVE diminished survival and proliferation while stimulating apoptosis in human breast cancer (BC) MCF-7 cells12,13. Anethole, a major component of star anise, exhibited promising anticancer effects against estrogen receptor-positive (MCF-7) and estrogen receptor-negative (MDA-MB-231) BC cells via upregulating Caspase-9, c-FLIP, and P5314. Furthermore, trans-anethole (1-methoxy-4-[1(E)-propenyl] benzene) suppressed osteosarcoma cells by generating reactive oxygen species (ROS) and arresting the cell cycle at the G0/G1 phase. It also induced the intrinsic apoptosis pathway, associated with increased levels of P53 and Caspases 3 and 915. Although the apoptotic effects of IVE and some of its major constituents, such as anethole, have been reported in different cancer models, the combined effects of IVE with PTX in prostate cancer cells—particularly in androgen-independent PC3 and androgen-sensitive LNCaP cells—remain insufficiently investigated. Therefore, the present exploratory in vitro study aimed to evaluate whether IVE could enhance PTX-associated antiproliferative and pro-apoptotic responses at the cellular and transcriptional levels. Importantly, this research was designed as a preliminary screening investigation rather than a formal pharmacological synergy analysis. Specifically, we sought to explore whether IVE could enhance PTX-induced cytotoxicity in PC3 and LNCaP cells at the cellular and transcriptional levels within an experimental in vitro framework.
Materials and methods
The present in vitro experiment followed the guidelines of the Ethics Committee of the Kermanshah University of Medical Sciences, Kermanshah, Iran [Ethics no.: IR.KUMS.MED.REC.1403.025].
IVE preparation
Illicium verum Hook f. (fruits and seeds) were purchased from the local herbal market, and authenticated by a botanist. One hundred grams of dried Star anise fruit and seed were powdered, and dissolved in 500 ml of 70% ethanol for 48 h under gentle shaking at room temperature (percolation method). Then, the prepared extract was filtered, poured into a Petri dish, and left in a safety cabinet to evaporate the ethanol at room temperature. The obtained extract was collected and stored at 4°C. Finally, for cell culture experiments, the extract powder was dissolved in the serum-free RPMI medium and filtered through a 0.22 μm membrane filter. Whole experiments were administered using a single extract batch to minimize batch-to-batch variability. The extraction yield was recorded for the prepared extract. However, comprehensive phytochemical characterization of the crude hydroalcoholic extract, such as HPLC, GC-MS, or LC-MS profiling, was not performed in the present study. Therefore, the exact composition and concentration of major bioactive constituents, including anethole, were not quantified. All cell culture experiments were performed using a single extract batch to minimize batch-to-batch variability. The lack of phytochemical standardization is acknowledged as a limitation, and the findings should be interpreted as preliminary screening results.
Cell line preparation and culture
Human prostate adenocarcinoma PC3 and LNCaP cell lines were obtained from the Pasteur Institute of Iran (Tehran, Iran). Cells were cultured in RPMI-1640 medium (Gibco, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, USA), 100 U/mL penicillin, and 100 µg/mL streptomycin (Sigma-Aldrich, Merck, Germany). Cells were maintained at 37°C in a humidified incubator containing 5% CO2. The culture medium was replaced every other day until the cells reached approximately 80% confluence. Paclitaxel (PTX, ≥98% purity) was purchased from Sigma-Aldrich (Merck, Germany).
After PC3 and LNCaP cells reached the desirable confluences, they were detached from flasks using a 0.25% trypsin/EDTA solution and centrifuged at 1800 rpm for 10 minutes. The cell pellets were re suspended in the complete medium and cultivated at the bottom of 96-well plates at 1×10 cells/well density. The cells were treated with different concentrations of PTX (0, 5, 10, 25, 50, and 100 nM) and IVE (0, 12.5, 25, 50, and 100 µg/ml) for 24, 48, and 72 h.
Cell proliferation and IC calculation
An MTT (3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide) assay was performed to evaluate the growth inhibitory effects of IVE and PTX. Each time, culture media were discarded from wells and replaced with 100 µl MTT solution (0.5 mg/ml), followed by incubating at 37°C for 3 h. The MTT reagent was then replaced with 100 µl DMSO to solve formazan crystals. The samples' optical density (OD) was measured at 570 nm using an ELISA reader (STATFAX2100, Biorad, Hercules, CA, USA). Cell proliferation was calculated according to the equation (1): Cell proliferation (%) = (OD treated sample / OD control sample) × 100
In the equation, OD is the OD of treated cells, and OD is the OD of control (untreated) cells. The half-maximal inhibitory concentration (IC) values were calculated using the dose-response curve plotted in GraphPad Prism software. IC values were expressed as concentrations, namely nM for PTX and µg/mL for IVE. Cell proliferation values were expressed as percentages and were not used interchangeably with IC values.
Real-time PCR assay
The effect of PTX, IVE, and PTX-IVE treatment (Combination treatments were performed using the IC concentrations of PTX and IVE determined at 48 h. For combination treatment, cells were simultaneously exposed to the 48 h IC concentration of PTX and the 48 h IC concentration of IVE for each cell line. The expression of BAX, BCL-2, P53, Caspase-3, and Caspase-9 genes were evaluated using the real-time PCR. After PC3 and LNCaP cells were treated with IC values of each PTX, IVE, and PTX-IVE for 48 h, cells were collected from plates, and total RNA was extracted using the Trizol reagent (Life Biolab, Heidelberg, Germany). RNA purity was assessed Nanodrop system (NanoDrop 2000, Thermo Scientific, Germany, Deutschland), and samples with acceptable A260/280 ratios were used for cDNA synthesis. 1 µg of RNA was utilized for cDNA synthesis using a commercial kit (ThermoFisher Scientific Inc., Porto Salvo, Portugal). Real-time PCR was carried out by using cDNA, a high-ROX SYBR green PCR Master Mix (RealQ Plus 2x Master Mix, Ampliqon, Denmark) and specific primers in the Step One Real-time PCR machine (Applied Biosystems, Waltham, Massachusetts, USA)1.
Experiments were performed in triplicate to ensure reproducibility. Primer specificity was confirmed by melt curve analysis, and no nonspecific amplification or primer-dimer formation was observed. The comparative threshold cycle (C) values were normalized against the housekeeping glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene, and relative gene expression was reported as the Fold change = 2. Forward and reverse primers are listed in the following Table 1.
Forward and Reverse Primers Sequences used for Real time PCR
| Tm (°c) | Length (bp) | Sequence (5'→3') | Accession no. | Gene |
|---|---|---|---|---|
| 60 |
24 24 |
F: cctgtgcaccaaggtgccggaact R: ccaccctggtcttggatccagccc | NM_001291430.2 | |
| 60 |
24 24 |
F: ttgtggccttctttgagttcggtg R: ggtgccggttcaggtactcagtca | NM_000657.3 | |
| 60 |
19 19 |
F: tggactgtggcattgagac R: caaagcgactggatgaacc | NM_032991.3 | |
| 60 |
22 22 |
F: gtttgaggaccttcgaccagct R: caacgtaccaggagccactctt | NM_001229 | |
| 60 |
21 22 |
F: taacagttctgcatgggcggc R: aggacaggcacaaacacgcacc | NM_001126118.2 | |
| 60 |
22 22 |
F: aaggtcggagtcaacggatttg R: gccatgggtggaatcatattgg | NM_001289745.3 |
Statistical analysis
Quantitative data were collected in triplicates (n=3) and reported as Mean ±SD. Data were analyzed using a One-way analysis of variance (ANOVA) followed by Tukey's test in GraphPad Prism 7 software (GraphPad Software, San Diego, CA, USA) at the significant level of P < 0.05.
Results
Cell proliferation
The effect of various concentrations of PTX on the proliferation of PC3 and LNCaP cells was evaluated using the MTT assay (Figure 1A-B). The effect of various concentrations of IVE on the proliferation of PC3 and LNCaP cells was evaluated using the MTT assay (Figure 2A-B). PTX and IVE decreased cell proliferation in a time and dose-dependent manner (*** P ˂ 0.001, ** P ˂ 0.01, * P ˂ 0.05). As illustrated in Figure 1, treatment with PTX resulted in a significant time- and dose-dependent reduction in the proliferation of both PC3 and LNCaP cell lines. The IC values for PTX in PC3 cells were determined to be 19.2 nM, 12.5 nM, and 8.3 nM at 24, 48, and 72 h, respectively. In LNCaP cells, the corresponding IC values were 11.8 nM, 8.1 nM, and 8.3 nM for the same time points. At the highest tested concentration (100 nM), cell viability dropped to minimum values of 23.2 ± 0.7, 19.5 ± 1.0%, and 13.8 ± 1.5% for PC3 cells, and 19.5 ± 0.8%, 13.6 ± 1.2%, and 9.6 ± 1.0% for LNCaP cells at 24, 48, and 72 h, respectively. At the highest tested IVE concentration, cell proliferation decreased to 58.5 ± 1.6%, 31.2 ± 1.7%, and 26.6 ± 4.5% in PC3 cells and to 29.5 ± 2.2%, 25.6 ± 1.6%, and 23.3 ± 2.6% in LNCaP cells after 24, 48, and 72 h, respectively. (Figure 2A-B). IC values (mean ± SD) of PTX and IVE in PC3 and LNCaP prostate cancer cells at different time points, as determined by MTT assay in Table 2. Significant differences in cell proliferation were either observed between untreated cells (control) with treated cells and among treated cells (*** P ˂ 0.001, ** P ˂ 0.01, * P ˂ 0.05). IC values of PTX and IVE were calculated for each cell line (Figure 3A-B).
IC50 values of PTX and IVE in prostate cancer cell lines
| Cell Line | Treatment | IC50 (µg/mLor nM) – 24h | IC50 (µg/Ml or nM) – 48h | IC50(µg/ml or nM)- 72h |
|---|---|---|---|---|
| PC3 | PTX | 23.2 ± 0.7% | 19.5 ± 1% | 13.8 ± 1.5% |
| PC3 | 58.5 ± 1.6% | 31.2 ± 1.7% | 26.6 ± 4.5% | |
| LNCaP | PTX | 19.5 ± 0.75% | 13.6 ± 1.15% | 9.6 ± 1% |
| LNCaP | 29.5 ± 2.2% | 25.6 ± 1.6% | 23.3 ± 2.6% | |
| Cell Line | Synergic treatment ( | |||
| PC3 | 31.2 ± 1.7% µg/mL + 19.5 ± 1% nM | |||
| LNCaP | 25.6 ± 1.6% µg/mL + 13.6 ± 1.15% nM | |||
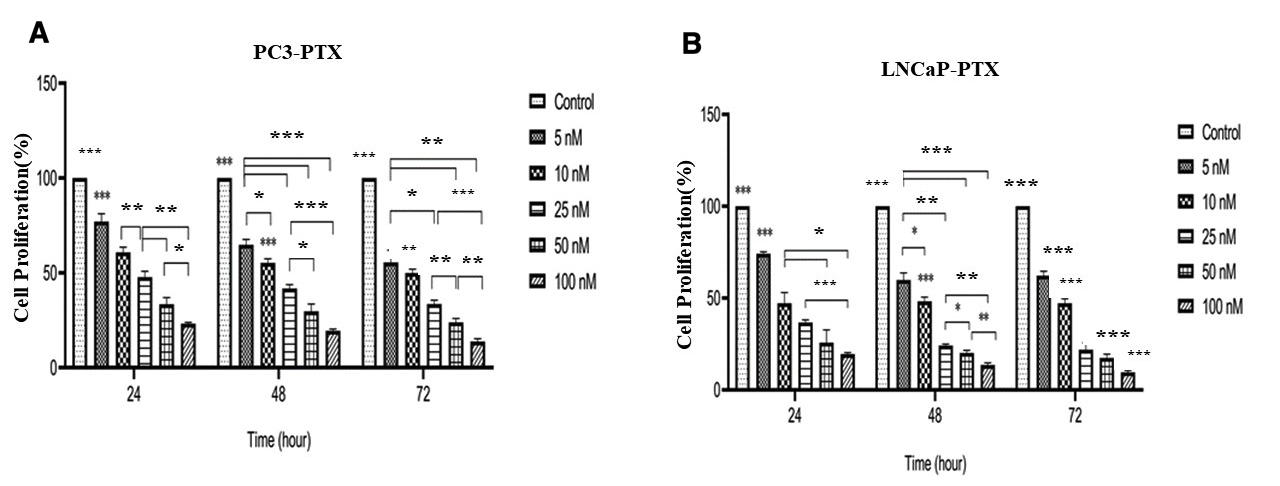
Inhibitory effect of various concentrations of PTX on the proliferation of PC3 (A) and LNCaP (B) cells, as assessed by the MTT assay. Cells were treated with PTX for 24, 48, and 72 h. Data are presented as mean ± SD from three independent experiments. *P < 0.05, P < 0.01, and ***P < 0.001 compared with the untreated control group.
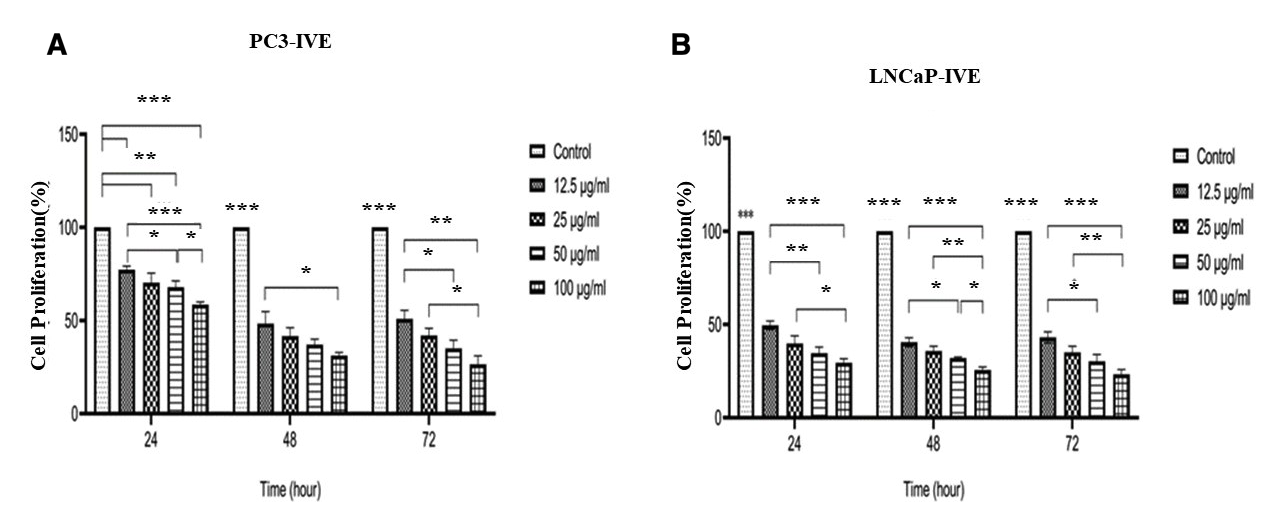
Inhibitory effect of various concentrations of IVE on the proliferation of PC3 (A) and LNCaP (B) cells, as assessed by the MTT assay. Cells were treated with IVE for 24, 48, and 72 h. Data are presented as mean ± SD from three independent experiments. *P < 0.05, P < 0.01, and ***P < 0.001 compared with the untreated control group.
The estimated IC values of PTX for PC3 cells were 19.2, 12.5, and 8.3 nM at 24, 48, and 72 h. For LNCaP cells, the corresponding IC values were 11.8, 8.1, and 8.3 nM. (Figure 3A-B). The estimated IC value of IVE in PC3 cells could not be calculated at 24 h because a clear dose-dependent inhibitory response sufficient for IC estimation was not obtained. The IC values of IVE in PC3 cells were 11.8 and 14.1 µg/mL at 48 and 72 h. In LNCaP cells, the IC values of IVE were 12.23, 7.1, and 8.8 µg/mL at 24, 48, and 72 h. (Figure 4A-B).
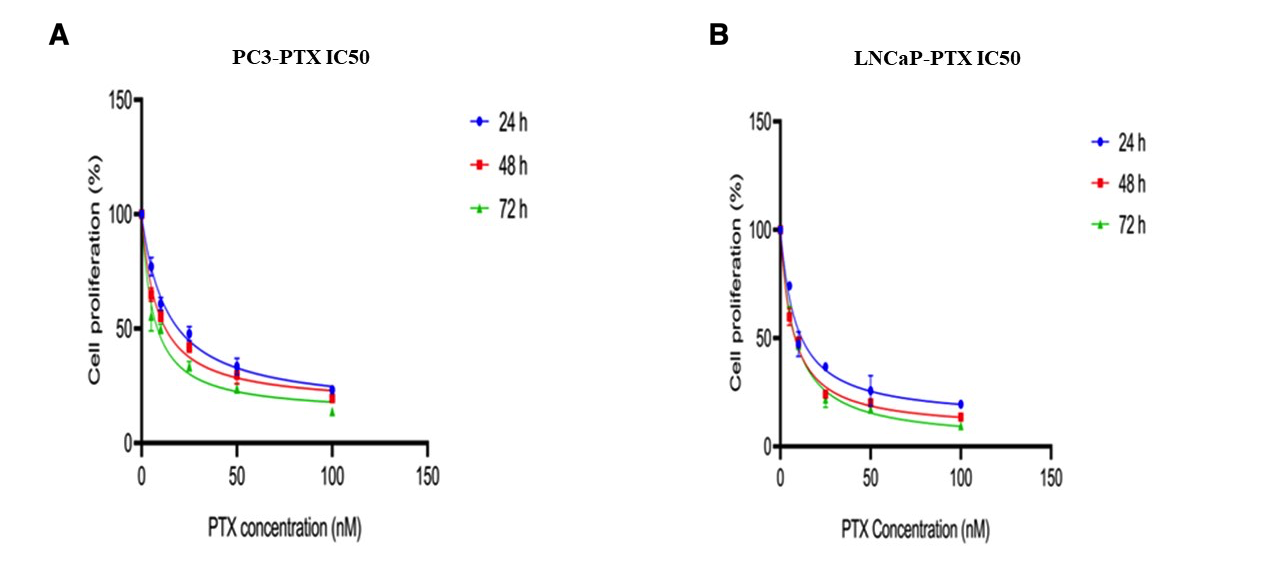
Dose–response curves and estimated IC50 values of PTX in PC3 (A) and LNCaP (B) cells after 24, 48, and 72 h of treatment.
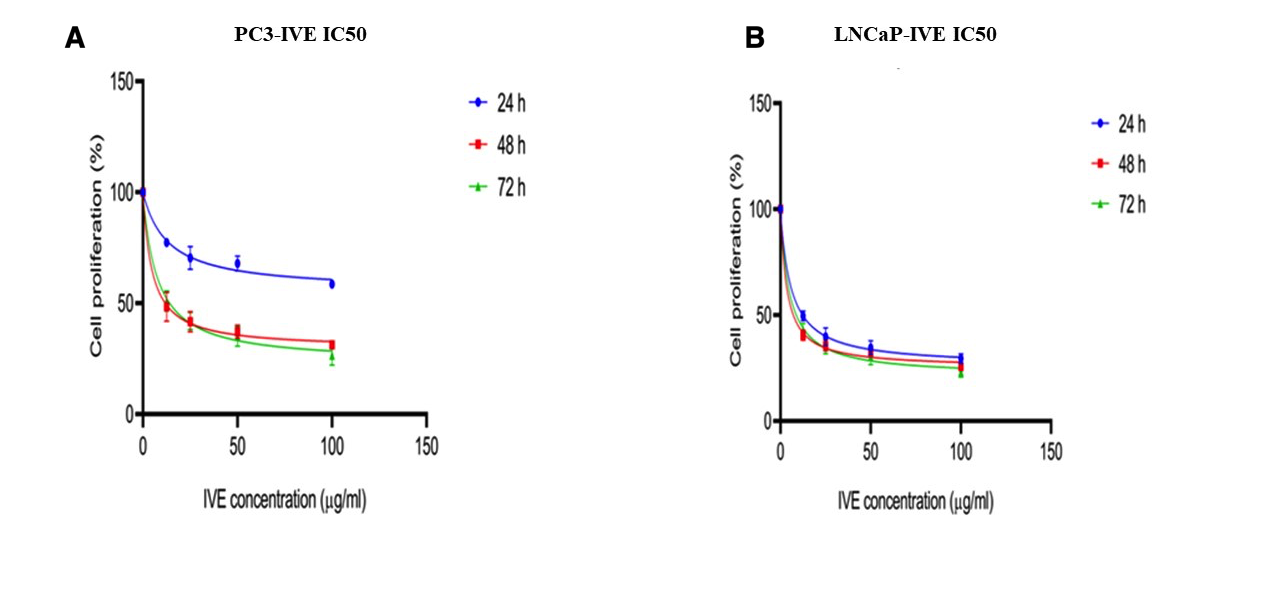
Dose–response curves and estimated IC50 values of IVE in PC3 (A) and LNCaP (B) cells after 24, 48, and 72 h of treatment.
Gene expression
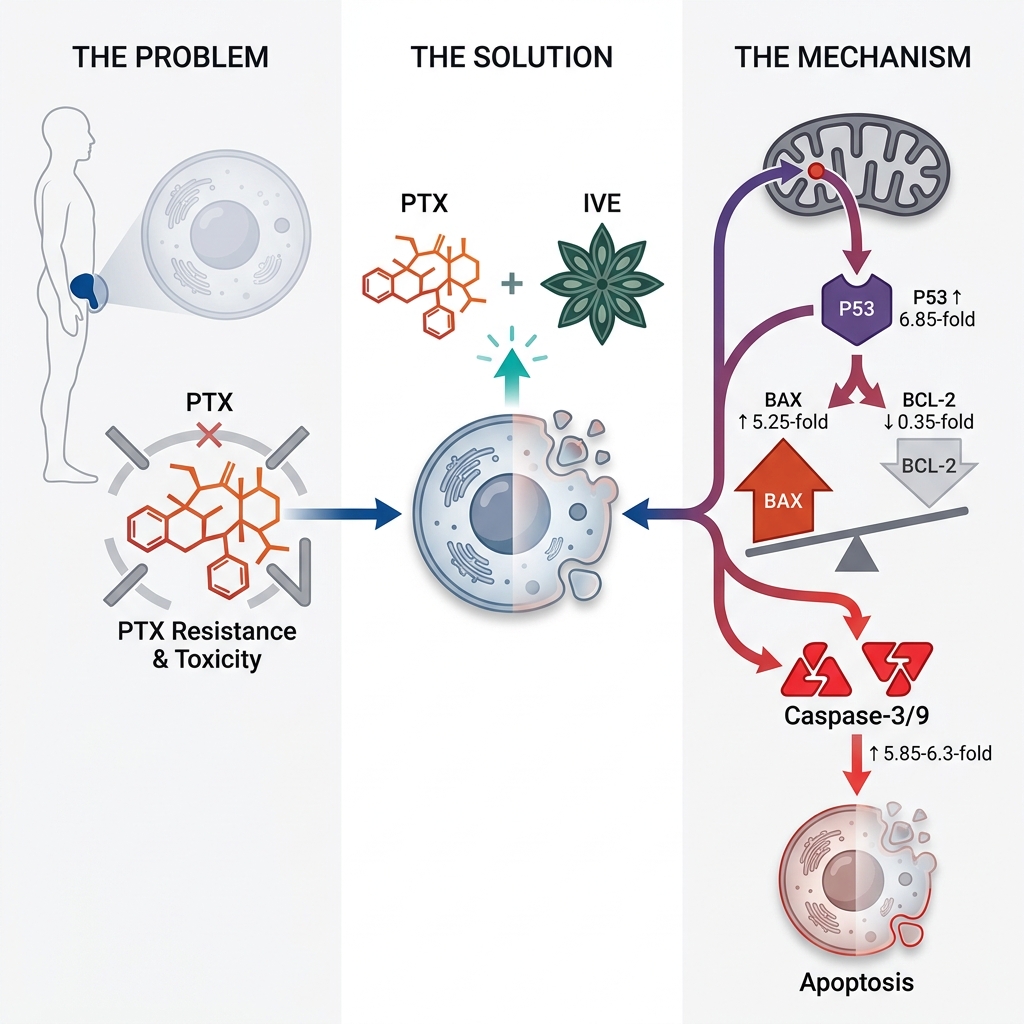
The mRNA expression levels of P53, BAX, BCL-2, Caspase-3, and Caspase-9 were evaluated in PC3 and LNCaP cells treated for 48 h with PTX, IVE, or their combination at the corresponding 48 h IC concentrations. (Figure 5, 6). Significant differences in gene expression were observed between the control group and treatment groups (*** P ˂ 0.001). For PC3 cells, P53 expression was 2.4 ± 0.1, 3.4 ± 0.2, and 6.85 ± 0.02 times greater in PC3 cells treated with PTX, IVE, and PTX-IVE, with remarkable differences and the highest value in the combination treatment group (Figure 5A) (*** P ˂ 0.001). BAX expression in PC3 cells subjected to PTX, IVE, and PTX-IVE was 2.33 ± 0.06, 2 ± 0.06 and 5.25 ± 0.25 times more than the control cells and the strongest impact was observed in PTX-IVE co-treatment group (Figure 5B) (*** P ˂ 0.001, ns: non-significant).
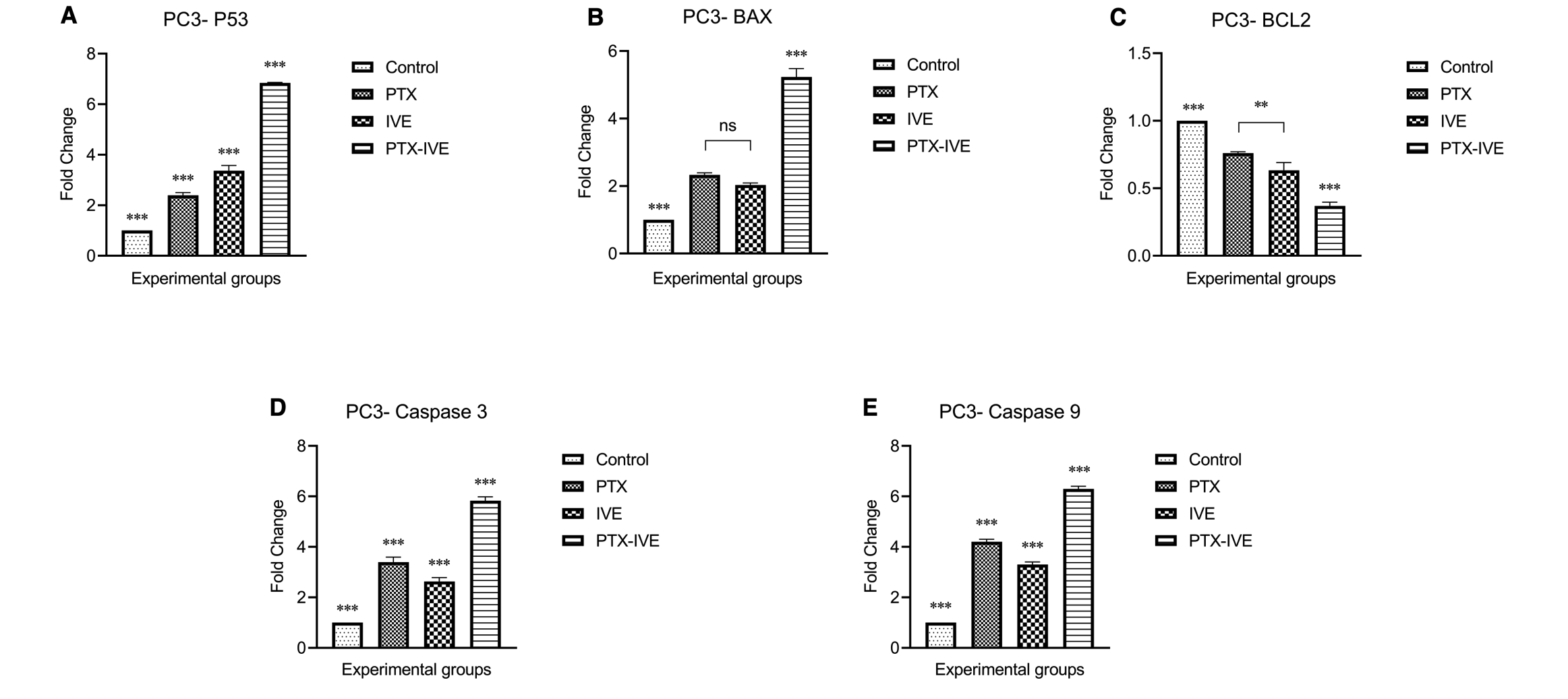
Relative mRNA expression levels of
Relative mRNA expression levels of
BCL-2 decreased significantly in PTX, IVE, and PTX-IVE co-treatment groups to 0.76 ± 0.01, 0.65 ± 0.06, and 0.35 ± 0.03 folds and the highest decline was detected in PTX-IVE combination (Figure 5C) (*** P ˂ 0.001, ** P ˂ 0.01). Caspase-3 levels were higher in treated PC3 cells with 3.4 ± 0.2, 2.65 ± 0.15, and 5.85 ± 0.15 values in PTX, IVE, PTX-IVE co-treatment groups and the highest values was found in the PTX-IVE co-treatment group (Figure 5D) (*** P ˂ 0.001). Caspase-9 values in PTX, IVE, and PTX-IVE co-treatment groups were remarkably higher with 4.2 ± 0.1, 3.3 ± 0.1, and 6.3 ± 0.1 folds and the most expression was observed in PTX-IVE co-treatment group (Figure 5E) (*** P ˂ 0.001).
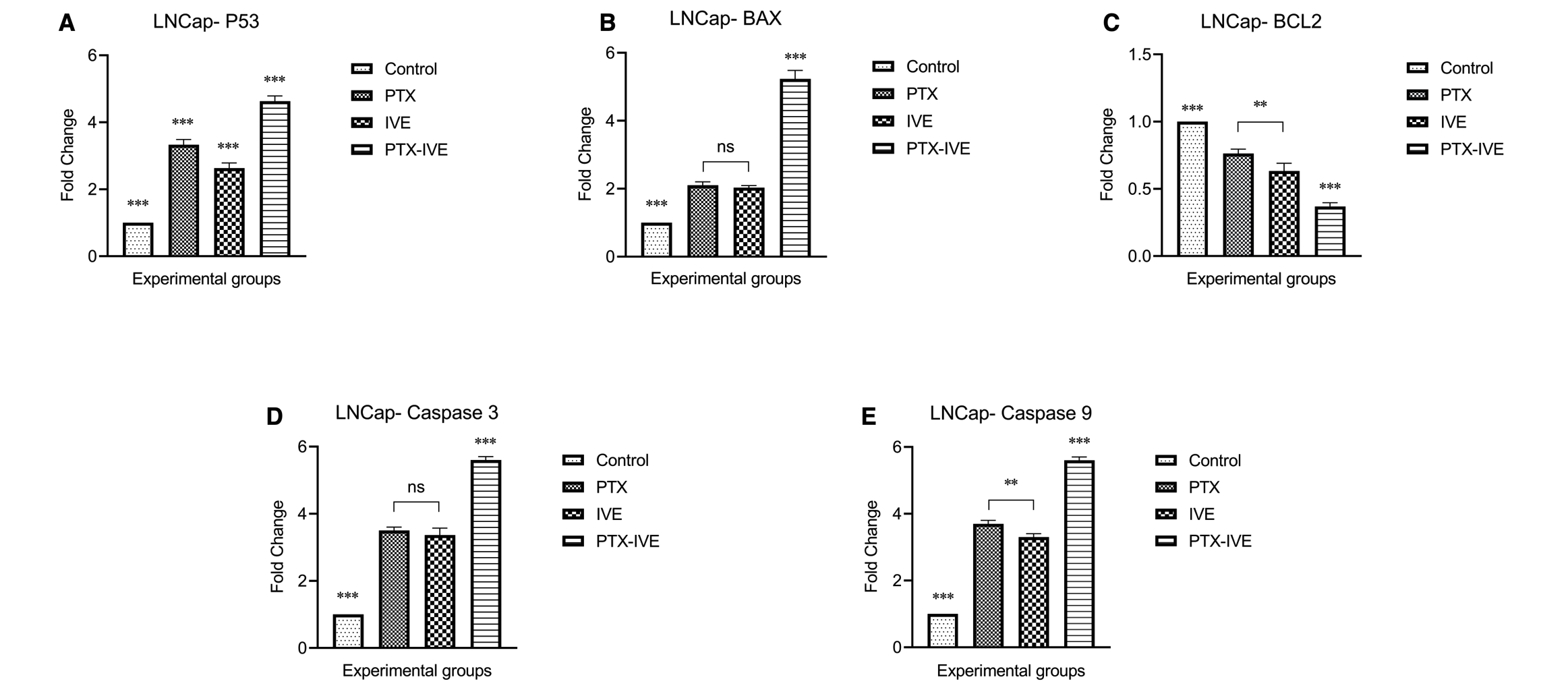
For LNCaP cells, P53 enhanced 3.35 ± 0.15, 2.65 ± 0.15, and 4.63 ± 0.15 folds in PTX, IVE, and PTX-IVE co-treatment groups compared with the control group with the highest value in PTX-IVE co-treatment group (Figure 6A) (*** P ˂ 0.001). BAX levels increased drastically in treatment groups with 2.1 ± 0.1, 2.05 ± 0.06, and 5.25 ± 0.25 values in PTX, IVE, and PTX-IVE co-treatment groups and the strongest effect was detected in the PTX–IVE co-treatment group (Figure 6B) (*** P ˂ 0.001, ns: non-significant). BCL-2 showed a prominent decline in treated groups with 0.76 ± 0.03, 0.65 ± 0.06, and 0.35 ± 0.04 values in PTX, IVE, and PTX-IVE co-treatment groups (Figure 6C) (*** P ˂ 0.001, ** P ˂ 0.01). The expression of Caspase-3 escalated significantly in treatments as determined by 3.5 ± 0.1, 3.4 ± 0.2, and 5.6 ± 0.1 folds in PTX, IVE, and PTX-IVE co-treatment groups with the highest expression in the latter group (Figure 6D) (*** P ˂ 0.001, ns: non-significant). Caspase-9 overexpressed in PTX, IVE, and PTX-IVE co-treatment groups with 3.7 ± 0.1, 3.3 ± 0.1, and 5.6 ± 0.1 folds higher than the control with the highest level in the combination group (Figure 6E) (*** P ˂ 0.001, ** P ˂ 0.01).
Discussion
In the present study, we investigated the antiproliferative and pro-apoptotic effects of PTX and IVE, both alone and in combination, in PC3 and LNCaP prostate cancer cell lines. Despite the established antitumor activity of PTX, its clinical use can be limited by adverse effects and the development of drug resistance. In recent years, an increasing number of studies have highlighted the application of medicinal plants and natural compounds as adjunctive treatments for various types of cancer, including prostate, breast, cervical, lung, and skin cancers17,18,19,20.
I. verum is a traditional Chinese medicinal plant with potent antioxidant, antimicrobial, antiviral, antifungal, anti‐inflammatory, anthelmintic, antinociceptive, and gastroprotective effects that has been less studied as an anticancer agent21. Our results indicated that IVE inhibited cell proliferation, with 48-hour IC values of 11.8 µg/mL and 7.1 µg/mL in PC3 and LNCaP cells, respectively. Importantly, the differences observed between the two prostate cancer cell lines may be attributed to fundamental variations in their biological characteristics. PC3 cells, as an androgen-independent prostate cancer model, commonly exhibit altered P53-related signaling, whereas LNCaP cells are androgen-sensitive. These differences may partially explain the variability in the cellular responses to the PTX and IVE treatments. Previous studies have reported varying IC values depending on the cancer type and degree of malignancy. For instance, an IVE IC dose of 2.8 ± 0.6 µg/mL reduced the viability of MDA-MB-231 triple-negative breast cancer (TNBC) cells and subsequently stimulated apoptosis22. Furthermore, I. verum essential oil, with an IC value of 50.34 ± 1.19 μg/mL, displayed significant toxicity against HCT116 colon cancer cells, which was associated with the dose-dependent suppression of cell migration and invasion, and the induction of apoptosis23. Yadav et al. showed that treating N-nitrosodiethylamine (NDEA)/phenobarbital (PB)-induced hepatocarcinogenesis in rats with star anise extract at the promotion and initiation stages of tumor growth reduced the tumor burden by lowering oxidative stress24.
Our findings demonstrated that IVE alone or in combination with PTX, enhanced the expression of antitumor genes such as P53 and BAX, and triggered the caspase cascade of the intrinsic apoptotic pathway, while simultaneously decreasing the expression of the anti-apoptotic BCL-2 gene. P53 is a tumor suppressor gene with a critical role in preventing cancer development by regulating various cellular processes such as the cell cycle, metabolism, DNA repair, and apoptosis25. BAX (BCL2L4, BCL-2-associated X protein) is a pro-apoptotic protein from the BCL-2 family with a crucial role in governing the intrinsic pathway of apoptosis26. BCL-2 is an anti-apoptotic member of the BCL-2 family, and its upregulation is linked to cancer cell survival, tumor maintenance, and progression27.
Apoptosis is a type of programmed cell death necessary for morphogenetic homeostasis during early development, as well as under pathological conditions28. Apoptosis occurs via intrinsic (activated by cellular stress) and extrinsic (triggered by death receptors) pathways, each of which ultimately activates death proteases known as caspases29. The dysregulation of apoptotic pathways results in the progression of several diseases, including cancer30. Phytochemicals have shown great potential in augmenting the anticancer effects of chemotherapeutics by upregulating apoptosis in glioblastoma and BC cells31,32.
Limited studies have evaluated the anticancer effects of I. verum concerning its underlying molecular mechanisms. Anethole, a major bioactive compound isolated from star anise, was shown to inhibit MCF-7 and MDA-MB-231 breast cancer cells via upregulating P53 and suppressing its downstream inhibitor, NF-κB. It also activated both the extrinsic and intrinsic apoptotic pathways via prompting the expression of Caspase-8 and -914. trans-Anethole (1-methoxy-4-[1(E)-propenyl] benzene) inhibited osteosarcoma cell growth at the G0/G1 phase by producing reactive oxygen species (ROS). This triggered the intrinsic apoptotic pathway, associated with P53, Caspase-3, and Caspase-9 overexpression12,33. Our experiment showed that the co-treatment of PC3 and LNCaP cells with PTX and IVE increased the expression of the BAX gene while decreasing BCL-2 gene expression. BAX overexpression sensitizes PC cells to apoptosis by targeting adaptive survival mechanisms, including store-operated calcium entry (SOCE) and endoplasmic reticulum calcium storage34.
Although the combined PTX–IVE treatment consistently manifested stronger antiproliferative responses relative to PTX or IVE alone, we cautiously describe this potential interaction between PTX and IVE, highlighting an enhanced antiproliferative and pro-apoptotic effect, while acknowledging that this study has several limitations. First, the present work was conducted as an in vitro screening experiment with a limited number of biological replicates (n = 3), which may potentially reduce the statistical power of the analyses. Second, formal quantification of synergy, such as the Chou–Talalay combination index, was not performed. Third, the apoptotic effect was evaluated at the gene expression level, and confirmatory protein-based assays (e.g., Western blotting or flow cytometry) were not included. Fourth, detailed phytochemical characterization of the extract (e.g., HPLC analysis) was beyond the scope of the current in vitro study. Previous studies showed that I. verum constituents, especially anethole, have been demonstrated to elicit oxidative stress and mitochondrial dysfunction35,36. In this study, our findings show the upregulation of P53, the BAX/BCL-2 ratio, and Caspase genes in PC cell cancers; however, unlike studies that included particular phytochemical profiling, future investigations are required to link these effects to specific active constituents.
Conclusion
In conclusion, our results demonstrate that IVE could be suggested as a potential supplementary option to enhance chemotherapy efficacy and reduce the requirement for high doses of chemotherapeutics. However, there is limited preclinical evidence regarding the underlying mechanisms through which I. verum suppresses cancer growth and progression, which should be investigated in future experiments. Nevertheless, our findings are consistent with previous studies on the IVE-induced apoptotic effects in other cancers. The lack of phytochemical standardization and formal synergy quantification represent limitations of the current study.
Abbreviations
ANOVA: Analysis of variance; BAX: BCL-2-associated X protein; BC: Breast cancer; cDNA: Complementary deoxyribonucleic acid; CT: Comparative threshold cycle; DMSO: Dimethyl sulfoxide; EDTA: Ethylenediaminetetraacetic acid; FBS: Fetal bovine serum; FDA: Food and Drug Administration; GAPDH: Glyceraldehyde 3-phosphate dehydrogenase; GC-MS: Gas chromatography-mass spectrometry; HPLC: High-performance liquid chromatography; IC: Half-maximal inhibitory concentration; IV: Illicium verum Hook. f.; IVE: Illicium verum extract; LC-MS: Liquid chromatography-mass spectrometry; MRI: Magnetic resonance imaging; mRNA: Messenger ribonucleic acid; MTT: 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide; NDEA: N-nitrosodiethylamine; OD: Optical density; PB: Phenobarbital; PC: Prostate cancer; PCR: Polymerase chain reaction; PET: Positron emission tomography; PSA: Prostate-specific antigen; PTX: Paclitaxel; qRT-PCR: Quantitative real-time polymerase chain reaction; RNA: Ribonucleic acid; ROS: Reactive oxygen species; SD: Standard deviation; SOCE: Store-operated calcium entry; TNBC: Triple-negative breast cancer.
Acknowledgments
The authors would like to show their gratitude to the Fertility and Infertility Research Center, Health Technology Institute, Kermanshah University of Medical Sciences, Kermanshah, Iran for providing facilities and equipment.
Author’s contributions
All authors read and approved the final manuscript.
Funding
This study was supported financially by the Kermanshah University of Medical Sciences, Kermanshah, Iran [grant number: ].
Availability of data and materials
Data and materials used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Declaration of generative AI and AI-assisted technologies in the writing process
The authors declare that they have not used generative AI (a type of artificial intelligence technology that can produce various types of content including text, imagery, audio and synthetic data).
Competing interests
The authors declare that they have no competing interests.