
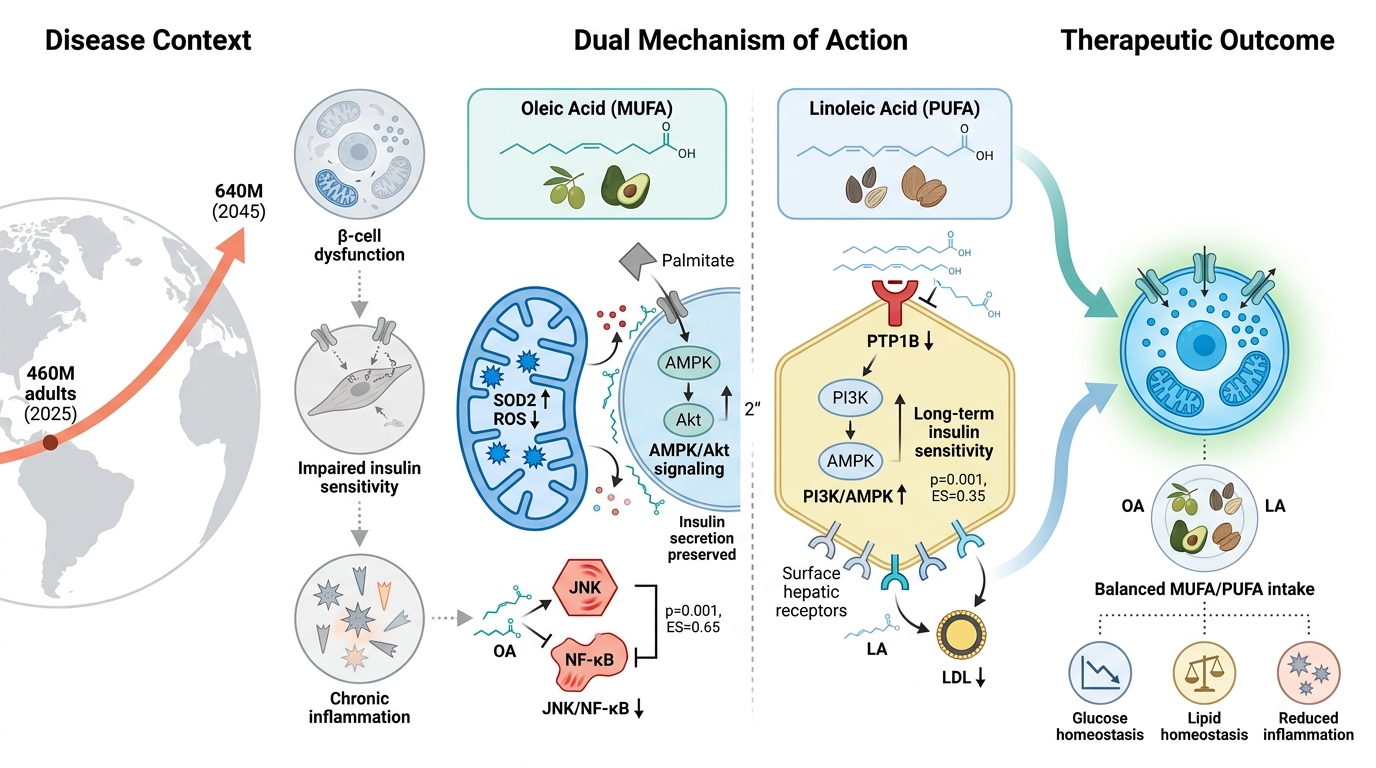

The Role of Monounsaturated and Polyunsaturated Fatty Acids: Oleic and Linoleic in Type 2 Diabetes Management
- Afe Babalola University Ado-Ekiti, Ekiti State, Km 8.5, Afe Babalola Way, Nigeria
- Department of Physiology, Olabisi Onabanjo University, Ago-Iwoye, Ogun State, Nigeria
Abstract
Dietary monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs)—particularly oleic acid (OA) and linoleic acid (LA)—are increasingly recognized as critical modulators of metabolic health and disease prevention. A growing body of evidence from clinical trials and experimental studies indicates that diets enriched in these unsaturated fats can improve insulin sensitivity, reduce systemic inflammation, and favorably alter lipid profiles compared to diets high in saturated fats. However, a limited number of studies report contradictory findings regarding the role of linoleic acid, particularly concerning lipid modulation and inflammation. Mechanistically, oleic and linoleic acids enhance insulin signaling through the activation of the PI3K/Akt/GLUT4 pathway, thereby promoting glucose uptake, while simultaneously suppressing pro-inflammatory cascades such as NF-κB and JNK. These dual actions highlight their potential efficacy in mitigating metabolic dysfunction in type 2 diabetes mellitus (T2DM). Despite consistent findings supporting their beneficial effects on glycemic control, debate persists regarding optimal intake levels, dose–response relationships, and long-term outcomes. This review synthesizes current knowledge, outlines dietary sources and intake guidelines, and identifies critical gaps in the research. Overall, oleic and linoleic acids demonstrate significant metabolic and anti-inflammatory benefits, positioning them as promising dietary components for T2DM management. However, standardized study designs and longitudinal investigations are required to clarify their translational relevance and establish evidence-based recommendations for clinical practice.
Introduction
Type 2 diabetes mellitus (T2DM) is among the fastest-growing noncommunicable diseases worldwide, driven by genetic predisposition, sedentary lifestyles, unhealthy diets, and obesity-related insulin resistance 1. More than 460 million adults currently live with diabetes, and projections suggest that this figure will exceed 640 million by 2045, underscoring the substantial epidemiological and economic burden of the disease (International Diabetes Federation, IDF Diabetes Atlas, 10th edition, 2021) 2. Pathophysiologically, T2DM is characterized by impaired insulin sensitivity, progressive β-cell dysfunction, chronic low-grade inflammation, and metabolic inflexibility, leading to dysregulated glucose and lipid homeostasis 3. These disturbances extend beyond glycemic control, contributing to endothelial dysfunction, hepatic steatosis, dyslipidemia, and heightened cardiovascular risk 4,5.
Dietary composition serves as a central determinant of metabolic health. Beyond mere caloric intake, the quality of dietary fats strongly influences insulin sensitivity, lipid metabolism, inflammatory processes, and cardiometabolic outcomes 6. Saturated fatty acids (SFAs) are consistently linked to lipotoxicity, inflammation, and impaired insulin signaling, whereas unsaturated fatty acids—particularly monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs)—exert beneficial metabolic effects 7. These benefits include improved membrane fluidity, enhanced lipid oxidation, the precise modulation of inflammatory pathways, and favorable changes in circulating lipoproteins 8. Among the unsaturated fats, oleic acid (OA), the predominant MUFA, and linoleic acid (LA), the major dietary ω-6 PUFA, are of particular clinical interest 9. OA supports membrane fluidity, activates the PPARα/γ pathways, and suppresses pro-inflammatory signaling cascades such as TLR4–NF-κB 10. Furthermore, LA contributes to membrane structure and generates bioactive metabolites that can exert either pro- or anti-inflammatory effects depending on the specific enzymatic pathways and tissue context 11.
This review synthesizes current evidence regarding the metabolic roles of OA and LA in T2DM management, highlighting mechanistic insights, findings from clinical and experimental studies, dietary sources, intake recommendations, and future research directions. Unlike earlier reviews that have examined MUFAs and PUFAs collectively, this work provides a direct mechanistic and clinical comparison between OA and LA, thereby clarifying their distinct yet complementary contributions to diabetes care.
Common Dietary Sources and approximate fatty acid content of Oleic Acid (MUFA) and Linoleic Acid (PUFA)
| Food Source | Oleic Acid (% of Total Fat) | Linoleic Acid (% of Total Fat) | Typical Serving Size | Calories per Serving (kcal) | Predominant Fatty Acid |
|---|---|---|---|---|---|
| Olive oil (extra virgin) | 70–75 | 8–11 | 1 tbsp (13.5 g) | 119 | Oleic acid (MUFA) |
| Avocado oil | 71–74 | 13–14 | 1 tbsp (14 g) | 124 | Oleic acid (MUFA) |
| Avocado (raw) | 41–58 | 8–15 | 100 g | 160 | Oleic acid (MUFA) |
| Canola oil | 60–64 | 18–28 | 1 tbsp (14 g) | 124 | Oleic acid (MUFA) |
| Almonds | 70–73 | 18–20 | 28 g (1 oz) | 164 | Oleic acid (MUFA) |
| Hazelnuts | 75–78 | 10–12 | 28 g (1 oz) | 178 | Oleic acid (MUFA) |
| Peanuts | 46–50 | 30–33 | 28 g (1 oz) | 161 | Oleic acid (MUFA) |
| Peanut oil | 48–49 | 32–34 | 1 tbsp (14 g) | 124 | Oleic acid (MUFA) |
| Sesame oil | 41–42 | 43–44 | 1 tbsp (14 g) | 120 | Mixed MUFA/PUFA |
| Walnuts | 22–23 | 52–63 | 28 g (1 oz) | 185 | Linoleic acid (PUFA) |
| Walnut oil | 23 | 63 | 1 tbsp (14 g) | 120 | Linoleic acid (PUFA) |
| Sunflower oil (conventional) | 20–30 | 59–69 | 1 tbsp (14 g) | 124 | Linoleic acid (PUFA) |
| Sunflower seeds | 20–25 | 55–65 | 28 g (1 oz) | 164 | Linoleic acid (PUFA) |
| Soybean oil | 24 | 58–60 | 1 tbsp (14 g) | 124 | Linoleic acid (PUFA) |
| Corn oil | 29 | 57 | 1 tbsp (14 g) | 122 | Linoleic acid (PUFA) |
Methodology
This study was conducted as a narrative review aimed at critically evaluating the existing evidence regarding the roles of monounsaturated and polyunsaturated fatty acids—particularly oleic acid (OA) and linoleic acid (LA)—in the management of type 2 diabetes mellitus (T2DM). The review focused on the underlying mechanisms of action, preclinical evidence, and clinical findings relevant to glucose and lipid metabolism.
A comprehensive literature search was performed using the PubMed, Scopus, Google Scholar, and Web of Science databases 16. Studies published between 2000 and 2025 were prioritized, while seminal older works were included to provide foundational context 16. Search terms combined keywords and Medical Subject Headings (MeSH) terms, such as "oleic acid," "linoleic acid," "monounsaturated fatty acids," "polyunsaturated fatty acids," "type 2 diabetes mellitus," "insulin resistance," "mechanism of action," "preclinical studies," "clinical trials," "glycemic control," "lipid metabolism," and "dietary fatty acids.". Boolean operators (“AND” and “OR”) were applied to refine the literature retrieval process 17.
Eligibility criteria encompassed in vitro experiments, animal studies, preclinical investigations, randomized controlled trials, cohort and observational studies, as well as systematic reviews and meta-analyses evaluating OA and LA in relation to T2DM. Only full-text articles published in English were included. Exclusions comprised duplicates, conference abstracts lacking methodological detail, and studies falling outside the scope of the review 18. Screening was conducted at the title, abstract, and full-text levels.
Data extraction captured the study design, population or experimental model, intervention duration, dosage and source of fatty acids, molecular pathways investigated, metabolic outcomes assessed, and principal findings relevant to diabetes management 18.
Findings were narratively synthesized and organized thematically under the categories of mechanisms of action, preclinical evidence, and clinical evidence. This approach enabled an integrated discussion of current knowledge, therapeutic implications, and research gaps regarding the potential roles of OA and LA in the prevention and management of T2DM.
Mechanisms of MUFA (Oleic Acid) and PUFA (Linoleic Acid) in Type 2 Diabetes
Oleic acid (OA), the principal monounsaturated fatty acid (MUFA), and linoleic acid (LA), the predominant dietary ω-6 polyunsaturated fatty acid (PUFA), exert complementary effects on metabolic regulation in type 2 diabetes mellitus (T2DM), particularly in preclinical models 19. Evidence from cellular, animal, and human studies highlights their roles in insulin signaling, β-cell preservation, inflammation, lipid metabolism, oxidative stress, and immune modulation 20,21,22.
In preclinical models, OA enhances insulin sensitivity by activating AMPK and augmenting Akt phosphorylation in cardiomyocytes, endothelial cells, and smooth muscle cells. Mechanistically, OA prevents the serine phosphorylation of IRS1 induced by TNF-α or palmitate, thereby preserving insulin receptor signaling under lipotoxic conditions 23. In contrast, palmitate activates the JNK/NF-κB pathways and disrupts insulin transduction 24. In vitro, LA strengthens insulin action by inhibiting protein tyrosine phosphatase 1B (PTP1B), increasing Akt phosphorylation, and activating the PI3K/AMPK pathways 25. Unsaturated fatty acids, including OA and LA, also protect pancreatic β-cells against palmitate-induced apoptosis, thereby promoting glycogen synthesis and glucose transport 26.
Both fatty acids significantly influence β-cell function 25,26,27,28,29. OA stimulates insulin secretion, reduces oxidative and endoplasmic reticulum (ER) stress, and rescues β-cells from palmitate-induced apoptosis 25,26,27. LA likewise protects β-cells; however, chronically high intake—especially in combination with high-sugar diets—may impair insulin secretion via endocannabinoid pathways 28,29.
OA demonstrates robust anti-inflammatory actions by increasing membrane fluidity, displacing arachidonic acid, reducing pro-inflammatory eicosanoids, and suppressing JNK/NF-κB activation while elevating IL-10 and adiponectin levels 28,30. Unlike saturated fats, OA does not activate the NLRP3 inflammasome 30. LA exerts more complex effects: it can suppress adaptive immune responses but may enhance innate immune activation 31. Conversely, moderate LA intake generally improves adipose tissue function via PPAR-γ activation 32.
Both fatty acids improve lipid metabolism through distinct pathways 18,33. OA protects against intracellular lipotoxicity by channeling saturated fats into triglyceride pools and enhancing mitochondrial β-oxidation 34. LA improves systemic lipid profiles by lowering LDL cholesterol, upregulating hepatic LDL receptors, and enriching adipose tissue PUFA pools 35. OA also exerts antioxidant and cytoprotective effects, lowering reactive oxygen species (ROS) and stabilizing mitochondria 4,27. Conversely, LA is more prone to peroxidation, carrying theoretical risks at very high intakes; however, clinical evidence regarding its role in oxidative stress in humans remains limited 36,37.
Taken together, OA and LA act through overlapping but distinct mechanisms to improve insulin sensitivity, reduce lipotoxic stress, and protect against diabetes-related complications 18,21. Their complementary roles suggest that a balanced dietary intake of MUFA- and PUFA-rich foods provides synergistic benefits in T2DM management.
Current Research and Evidence
Evidence from Preclinical Models
Preclinical studies strongly support the antidiabetic effects of oleic acid (OA). Rodent models of obesity and diabetes demonstrate that dietary or pharmacological supplementation with OA improves metabolic outcomes by reducing insulin resistance and hepatic steatosis when compared to saturated fat diets 10,18,38. Mechanistic investigations reveal that nitro-oleic acid, a nitrated derivative, activates the Nrf2 and PPAR-γ pathways, thereby reducing hepatic inflammation and improving glucose tolerance; notably, these protective effects in adipose and muscle tissues are dependent on intact Nrf2 signaling 39. In streptozotocin-induced or high-glucose rodent models, OA has been shown to preserve β-cell mass and insulin content relative to saturated fat diets 18, while its derivative oleoylethanolamide (OEA) attenuates palmitate-induced β-cell death through the hydrolysis and release of free oleate, independent of GPR119 signaling 27,40. Collectively, OA and its metabolites counteract insulin resistance, inflammation, oxidative stress, and β-cell loss in experimental models of diabetes 28.
In contrast, linoleic acid (LA) demonstrates more variable effects. Some rodent studies report improved insulin resistance and glucose homeostasis, with conjugated LA enhancing the PI3K/Akt and AMPK signaling pathways 41. However, excessive ω-6 intake may exacerbate inflammation, increasing pro-inflammatory cytokines such as IL-6 and impairing glucose control in obese diabetic rats maintained on high ω-6:ω-3 ratios 8,42. Direct comparisons between OA and LA remain limited; although both confer metabolic benefits relative to saturated fat, the optimal balance of ω-6 PUFA and ω-9 MUFA intake remains unresolved.
Evidence from Clinical Studies
Clinical investigations provide nuanced insights into the metabolic effects of OA and LA in T2DM. Epidemiological studies link higher OA intake, particularly from olive oil, with improved glycemic control and a reduced incidence of T2DM. Mediterranean diet interventions rich in OA lower fasting glucose, HbA1c, and insulin resistance compared with high-saturated-fat diets; furthermore, clinical trials further demonstrate improved endothelial function, reduced oxidative stress, and the preservation of β-cell function, closely aligning with preclinical findings 6,43,44.
Human evidence regarding LA is more complex. Large cohort studies and meta-analyses associate higher LA intake and circulating biomarkers with a reduced risk of T2DM, and Mendelian randomization supports a causal link between LA levels and improved glycemic traits, such as fasting glucose and insulin sensitivity 45. Clinical trials show that LA-rich diets increase adiponectin levels and improve lipid profiles, thereby indirectly supporting insulin sensitivity. However, excessive LA intake, particularly in the context of high ω-6:ω-3 ratios, may promote metabolic inflammation, underscoring the importance of dietary balance 31,46,47.
Comparative Evidence
Direct head-to-head clinical comparisons between OA and LA remain scarce. Available data suggest that both fatty acids confer metabolic benefits relative to saturated fats while acting through distinct mechanisms: OA primarily enhances insulin signaling and antioxidant defenses, whereas LA improves lipid handling and adipokine regulation 18,27,31,48. Taken together, these findings indicate that OA and LA are complementary, supporting the rationale for a balanced intake of MUFA- and PUFA-rich foods in the dietary management of T2DM 21,33.
Insulin Signaling and Glucose Metabolism
Oleic acid (OA) directly enhances insulin signaling by activating AMPK and Akt, preserving IRS1 function under inflammatory or lipotoxic conditions, and supporting β-cell insulin secretion 27. Linoleic acid (LA) indirectly strengthens insulin action by inhibiting protein tyrosine phosphatase 1B (PTP1B), thereby amplifying Akt phosphorylation and the downstream PI3K/AMPK pathways 25. Both fatty acids protect β-cells from palmitate-induced apoptosis, although OA demonstrates more consistent, direct cytoprotective effects 33. Together, these MUFAs and PUFAs converge on insulin signaling pathways, thereby improving glucose uptake and metabolic resilience 18.
Inflammation and Immune Modulation
OA exerts robust anti-inflammatory effects by suppressing TNF-α-induced JNK/NF-κB activation, reducing IL-6 and TNF-α levels, and increasing IL-10 and adiponectin expression 27,30,49. Importantly, OA avoids the activation of the NLRP3 inflammasome, thereby preventing IL-1β release 27. LA demonstrates bidirectional effects: it can suppress adaptive immune responses by reducing Th1/Th17 differentiation and cytokine release, yet it may enhance innate immune activation in neutrophils and macrophages 8,42. Human dietary studies generally suggest that moderate LA intake does not exacerbate inflammation and may modestly improve adipose tissue function 48. Thus, MUFA provides consistent anti-inflammatory protection, while PUFA modulates immune responses in a context-dependent manner 48,50.
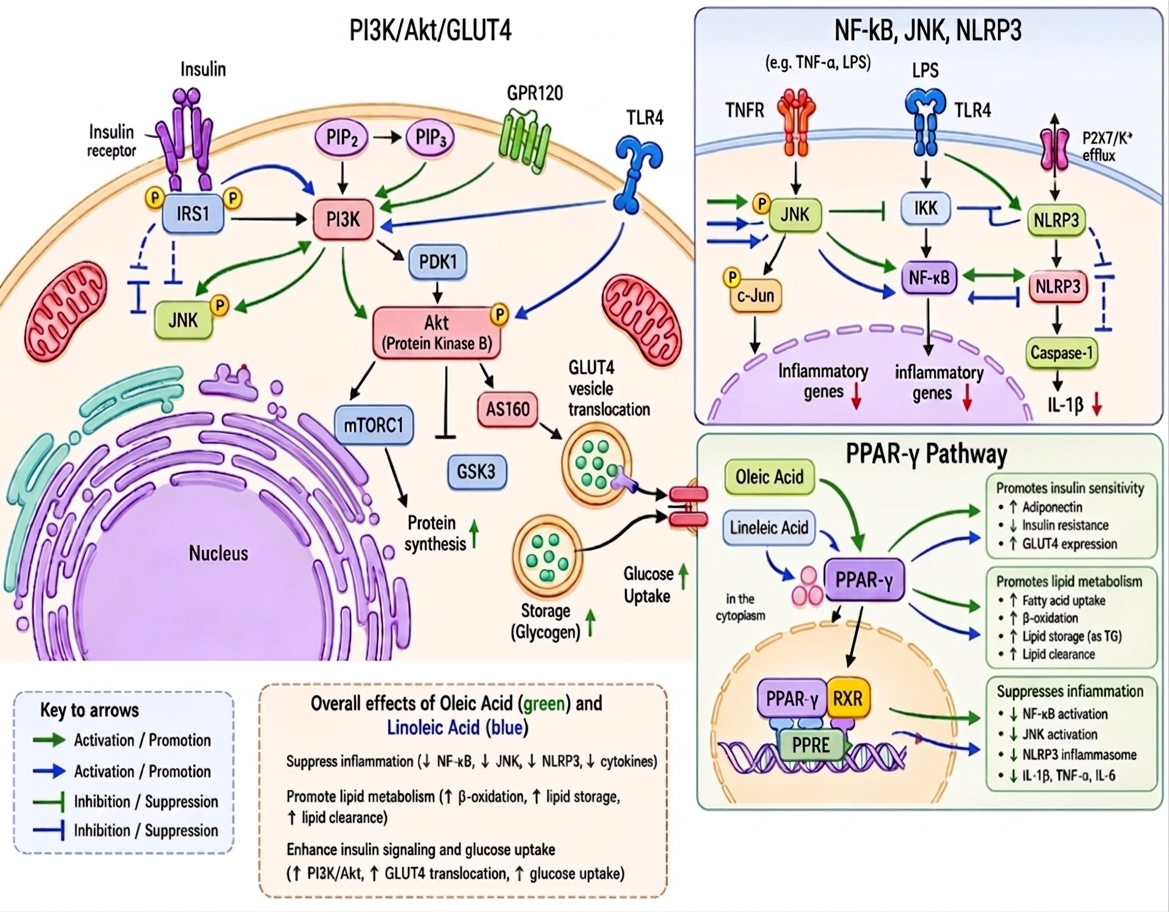
Mechanistic pathways of insulin signaling and inflammation: modulation by oleic and linoleic acids.
This figure illustrates the proposed molecular mechanisms of oleic acid (green arrows) and linoleic acid (blue arrows) in insulin signaling and inflammatory regulation. In the insulin signaling pathway, the activation of the insulin receptor and IRS1 stimulates PI3K, converting PIP2 to PIP3, which activates PDK1 and subsequently phosphorylates Akt 51,52. Akt promotes GLUT4 vesicle translocation and glucose uptake, while regulating GSK3, AS160, and mTORC1 27,35,52. OA enhances insulin sensitivity through GPR120 activation and the suppression of JNK, whereas LA modulates PI3K/Akt signaling and TLR4-associated pathways 3,40,41.
The inflammatory signaling panel depicts interactions among TNFR, TLR4, NF-κB, JNK, and the NLRP3 inflammasome 31,53,54. Both fatty acids attenuate inflammatory signaling by inhibiting JNK, NF-κB, and NLRP3 activation, thereby reducing downstream cytokine expression and IL-1β release 21,27,31. Excess LA intake may influence the TLR4/NF-κB and NLRP3 pathways in a context-dependent manner 8,42,55.
Beyond cytosolic crosstalk, OA and LA act as transcriptional ligands for PPAR-γ. Upon binding, PPAR-γ forms a heterodimer with RXR, translocates to the nucleus, and binds to peroxisome proliferator response elements (PPRE) 21,27,56. This complex enhances insulin sensitivity (via adiponectin and GLUT4 upregulation), accelerates lipid metabolism (fatty acid uptake, β-oxidation, and triglyceride storage), and suppresses chronic inflammation through the transrepression of NF-κB and JNK signaling, thereby reducing cytokine production (IL-1β, TNF-α, and IL-6). Collectively, these nuclear mechanisms demonstrate how OA and LA systematically diminish inflammatory stress, optimize lipid handling, and restore insulin sensitivity 21,27,56.
Lipid Metabolism
Oleic acid (OA) protects against intracellular lipotoxicity by channeling saturated fats into neutral triglyceride pools and enhancing mitochondrial β-oxidation, thereby preventing toxic lipid accumulation such as diacylglycerols (DAG) and ceramides 34. Linoleic acid (LA) improves systemic lipid profiles by lowering LDL and total cholesterol, upregulating hepatic LDL receptors, and enriching adipose tissue with PUFA-rich lipid pools 35. OA thus acts primarily at the cellular level, while LA exerts broader systemic lipid benefits 22.
Oxidative Stress and Cellular Protection
OA demonstrates strong antioxidant and cytoprotective properties, reducing reactive oxygen species (ROS), stabilizing mitochondrial function, and upregulating antioxidant defenses such as superoxide dismutase-2 (SOD2) 57. LA is more prone to peroxidation, yet moderate dietary intake does not significantly elevate oxidative stress biomarkers in humans 33. Its insulin-sensitizing and lipid-modulating effects generally outweigh potential oxidative risks 18,33. OA therefore provides more consistent direct antioxidant protection, whereas LA requires moderation to avoid peroxidative stress 57.
β-Cell Function
Both OA and LA preserve β-cell mass and function under lipotoxic conditions 18. OA enhances β-cell viability, reduces endoplasmic reticulum (ER) stress, and rescues cells from palmitate-induced apoptosis 28. LA protects β-cells via PI3K/Akt signaling, though excessive intake combined with high-sugar diets may impair insulin secretion through endocannabinoid pathways 29. Overall, OA offers more reliable β-cell protection, while LA’s effects are context-dependent 18.
Integrated Perspective
Oleic acid (OA, MUFA) and linoleic acid (LA, PUFA) act through overlapping yet distinct mechanisms to improve insulin sensitivity, reduce inflammation, and protect β-cells in T2DM 18,21. OA provides robust anti-inflammatory, antioxidant, and cytoprotective effects, while LA contributes to systemic lipid improvements and immune modulation 18,21,28. Their complementary actions suggest that balanced dietary intake of MUFA- and PUFA-rich foods—such as olive oil, nuts, and vegetable oils—may yield synergistic benefits in T2DM management 18,21.
Summary of key evidence: preclinical findings, clinical findings, mechanisms, dietary sources, and effect direction for oleic vs. linoleic acid
| Fatty acid | Outcome | Preclinical | Clinical/human | Mechanisms | Effect direction | References |
|---|---|---|---|---|---|---|
| Oleic acid | Insulin sensitivity | Preclinical | Oleic acid enhances insulin sensitivity by suppressing inflammatory kinases (JNK and IKKβ), preventing inhibitory IRS-1 serine phosphorylation, and preserving PI3K/Akt signaling. It reverses palmitate-induced insulin resistance through reduction of ROS generation, protection of mitochondrial function, and restoration of IRS-1/Akt signaling. Oleic acid also activates the cAMP/PKA/SIRT1–PGC-1α pathway to promote fatty acid oxidation, reduce DAG and ceramide accumulation, and maintain downstream insulin signaling under lipotoxic conditions. | favorable | ||
| Oleic acid | Insulin sensitivity | clinical | Oleic acid improves insulin sensitivity by activating the cAMP/PKA/SIRT1–PGC-1α pathway, enhancing fatty acid oxidation, and reducing intracellular DAG and ceramide accumulation, thereby preserving IRS/PI3K/Akt insulin signaling. Clinically, oleic acid-rich diets show superior glycemic effects compared with high-linoleic acid diets, producing lower fasting glucose, fasting insulin, and HOMA-IR levels in type 2 diabetes patients. Oleic acid also demonstrates protective interactions with the Pro12Ala polymorphism of the PPARG2 gene, significantly reducing diabetes risk. | favorable | ||
| Oleic acid | inflammation | preclinical | Oleic acid exerts potent anti-inflammatory effects by promoting anti-inflammatory M2 macrophage polarization while suppressing pro-inflammatory M1 activity. It inhibits NF-κB, MAPK, TLR4/NF-κB/NLRP3 inflammasome, and TNF-α signaling pathways, reducing inflammatory mediators including iNOS, COX-2, TNF-α, IL-6, IL-1β, IL-12, IL-6, and IL-8, while enhancing IL-10 expression and Treg cell survival. Oleic acid also sustains Akt/AMPK signaling through PP2A downregulation, upregulates FFAR4/GPR120, adiponectin, and PPAR-γ activity, and improves membrane lipid raft fluidity to prevent SFA-induced inflammatory kinase activation and adipose low-grade inflammation. | favorable | ||
| Oleic acid | inflammation | Clinical | Anti-Inflammatory Modulation: Up-regulates anti-inflammatory M2 macrophage polarization over pro-inflammatory M1 macrophages. It down-regulates Protein Phosphatase 2A (PP2A) to sustain Akt/AMPK activation, up-regulates free-fatty acid receptor-4 (FFAR4/GPR120), and increases insulin-sensitizing adiponectin expression while down-regulating pro-inflammatory TNF-α, IL-6, and IL-1β. Significantly dampens systemic circulating pro-inflammatory markers including TNF-α, IL-6, and C-reactive protein. | favorable | ||
| Oleic acid | Lipid profile | preclinical | Enhances the insulin signaling pathway and improves lipid metabolism. Reduces oxidative stress by decreasing reactive oxygen species (ROS) production and alleviates inflammation by inhibiting the NF-κB pathway and downregulating pro-inflammatory cytokines. Mitigates endoplasmic reticulum (ER) stress by downregulating stress markers, protecting beta cells from apoptosis | favorable | ||
| Oleic acid | Lipid profiles | Clinical | Oleic acid improves lipid metabolism and cardiovascular risk profiles by enhancing insulin signaling, reducing oxidative stress through decreased ROS production, and inhibiting NF-κB–mediated inflammatory responses. It mitigates endoplasmic reticulum (ER) stress, protecting β-cells from apoptosis and supporting metabolic homeostasis. Clinically, replacing saturated fatty acids with oleic acid–rich olive oil lowers LDL-C, triglycerides, total cholesterol, and fasting glucose while increasing HDL-C, with studies showing that ~30 mL/day olive oil intake for 4 weeks significantly improves lipid and glycemic markers in type 2 diabetes patients. Overall, oleic acid consistently improves serum triglycerides and optimizes LDL-C/HDL-C ratios when substituted for saturated fats. | favorable | ||
| Oleic acid | Oxidative stress | Preclinical | Prevents eNOS uncoupling, shifts cellular dynamics away from superoxide generation, and activates the Nrf2/HO-1 defense cascade. | favorable | ||
| Oleic acid | Oxidative stress | Clinical | Reduced NADPH oxidase activity and enhanced nitric oxide bioavailability in T2DM patients. | favorable | ||
| Oleic acid | β-Cell Function | Preclinical | Oleic acid protects pancreatic β-cells from glucolipotoxicity and apoptosis by reducing ROS production, upregulating mitochondrial SOD2, and inhibiting caspase-3/7–mediated cell death. It enhances cellular defense against SFA-induced lipotoxicity by promoting benign triglyceride droplet formation, preserving calcium-dependent insulin granule mobilization, and maintaining secretory function under metabolic stress. Oleic acid also alleviates ER stress and suppresses IL-6/IL-8 autocrine inflammatory loops, stimulates β-cell proliferation, and supports GLP-1 secretion from intestinal L-cells, thereby safeguarding β-cell integrity and improving overall insulin secretory capacity. | favorable | ||
| Oleic acid | β-Cell Function | clinical | Shields pancreatic islets from glucolipotoxicity and apoptosis by suppressing reactive oxygen species (ROS) production, upregulating mitochondrial superoxide dismutase 2 (SOD2), decreasing activated caspase-3 expression, and driving GLP-1 secretion from intestinal L-cells. | favorable | ||
| Linoleic acid | Insulin sensitivity | preclinical | Linoleic acid (including CLA) improves insulin sensitivity by activating key metabolic signaling pathways, notably PI3K-Akt and AMPK, through inhibition of negative regulators such as protein tyrosine phosphatases (PTPN1, PTPN9, PTPN11), thereby enhancing glucose uptake in adipocytes and peripheral tissues. In vivo studies show attenuation of high-fat diet–induced insulin resistance, fasting hyperglycemia, and glucose intolerance, alongside upregulation of insulin signaling and glucose transport in liver and adipose tissue. It also reduces ectopic fat deposition and systemic low-grade inflammation by downregulating pro-inflammatory transcriptional networks and improving immune–metabolic crosstalk, resulting in enhanced peripheral glucose utilization and reduced lipogenic signaling under balanced physiological conditions. | favorable | ||
| Linoleic acid | Insulin sensitivity | Clinical | Linoleic acid shows mixed short-term but beneficial long-term effects on insulin sensitivity. Although short-term high-LA diets may increase fasting glucose, insulin, and HOMA-IR compared with oleic acid diets, long-term studies consistently associate higher LA intake and circulating biomarkers with lower fasting glucose/insulin, improved HbA1c, enhanced peripheral insulin sensitivity, reduced OGTT glucose spikes, and lower type 2 diabetes risk. These benefits are linked to improved incorporation of LA into skeletal muscle and serum lipid fractions that support glucose utilization and insulin action. | favorable | ||
| Linoleic acid | Inflammation | preclinical | Linoleic acid exhibits dual inflammatory effects depending on metabolic context. Conjugated linoleic acid (CLA) demonstrates anti-inflammatory activity by blocking NF-κB nuclear translocation and reducing TNF-α, IL-6, IL-1β, and iNOS expression. However, excessive dietary linoleic acid can promote inflammation through its conversion to arachidonic acid, a precursor of pro-inflammatory eicosanoids generated via COX, LOX, and CYP450 pathways. High ω-6/ω-3 ratios further amplify the arachidonic acid cascade and increase pro-inflammatory oxylipin (OXLAM) production, contributing to chronic inflammatory disorders. | Mixed | ||
| Linoleic acid | Inflammation | Clinical | In human studies, linoleic acid demonstrates both beneficial and potentially pro-inflammatory effects. Long-term cohort studies show that higher circulating LA biomarkers are associated with significantly lower risk of type 2 diabetes (RR 0.65, 95% CI 0.60–0.72), with no adverse association observed for arachidonic acid. However, excessive LA intake may promote inflammation because LA is converted to arachidonic acid, a precursor of pro-inflammatory eicosanoids generated through COX, LOX, and CYP450 pathways, particularly under elevated ω-6/ω-3 dietary ratios. | Mixed | ||
| Linoleic acid | Lipid profile | Preclinical | Preclinical evidence shows that linoleic acid reduces liver triglyceride buildup by downregulating lipogenic genes and limiting abdominal fat, while polyphenols protect PUFAs from oxidative damage and enhance hepatic lipid transport. In parallel, gamma-linolenic acid (GLA) prevents lipid metabolism disorders by balancing autophagy and apoptosis via the LKB1–AMPK–mTOR pathway, boosting fatty acid oxidation through CPT1a and PPARα, lowering intracellular lipid content, and relieving oxidative stress by upregulating SOD and GPx and reducing ROS. Overall, PUFAs demonstrate protective effects on lipid metabolism and oxidative stress, though outcomes remain mixed and context-dependent, influenced by diet composition, fatty acid balance, and synergistic compounds. | Mixed | ||
| Linoleic acid | Lipid profile | Clinical | Dose–response meta-analyses of RCTs indicate that sustained high-dose CLA supplementation can adversely affect lipid profiles, elevating total cholesterol and LDL-C while lowering HDL-C. In contrast, standard dietary linoleic acid (LA) demonstrates robust hypocholesterolemic effects, reducing circulating total serum cholesterol and ApoB. Together, these findings highlight the divergent impacts of CLA and LA on lipid metabolism: CLA at high doses may impair cardiovascular risk markers, whereas LA intake supports healthier cholesterol regulation. | Mixed | ||
| Linoleic acid | β-Cell Function | preclinical | Prevents glucolipotoxicity-induced pancreatic beta-cell apoptosis. It safeguards islet cellular mass and maintains glucose-stimulated insulin secretion (GSIS) by counteracting endoplasmic reticulum (ER) stress triggered by exposure to high saturated fatty acid environments. | favorable | ||
| Linoleic acid | β-Cell Function | Clinical | Higher linoleic acid intake is associated with a 43% reduced risk of type 2 diabetes and lower fasting insulin levels. Mechanistically, it stimulates adipose tissue to secrete HMW adiponectin, a powerful insulin-sensitizing hormone. This systemic improvement lowers peripheral glucose load and insulin demand, indirectly shielding pancreatic beta-cells from chronic overstimulation, functional exhaustion, and glucolipotoxicity. | favorable |
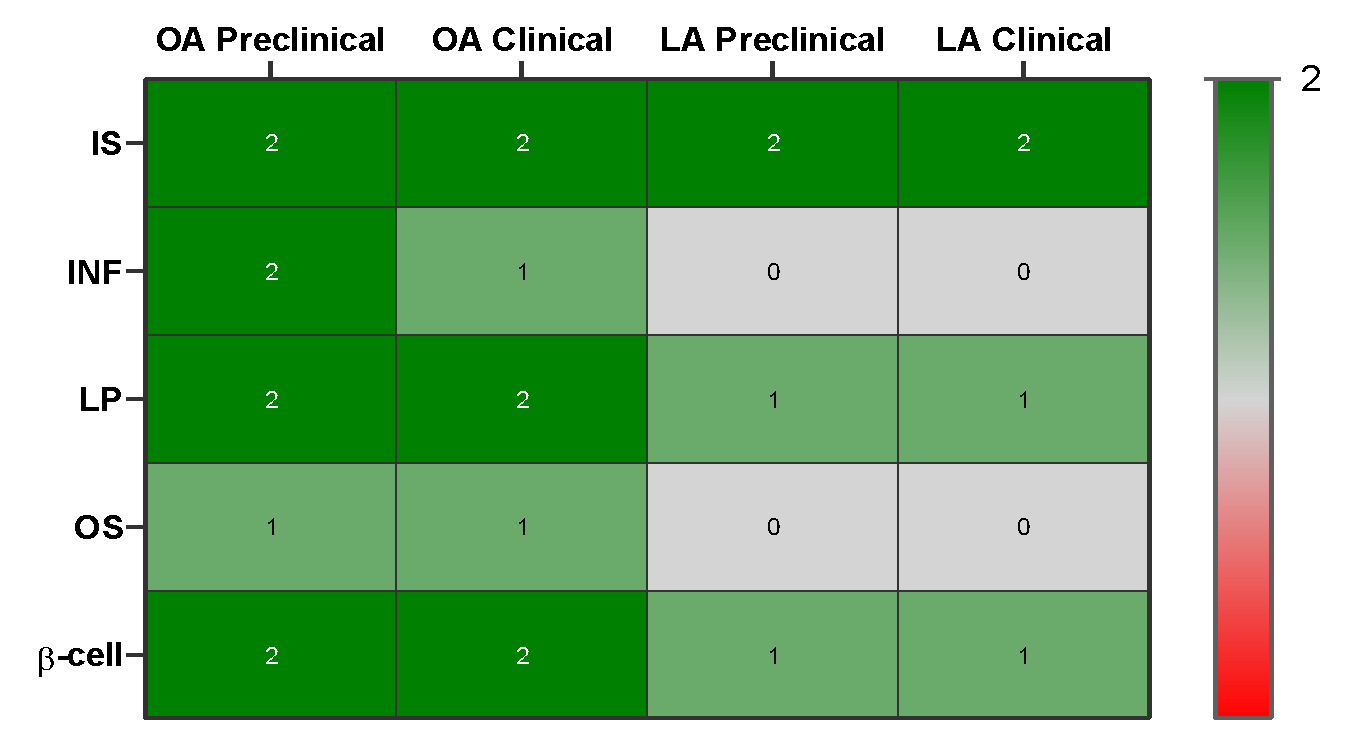
Evidence heatmap of qualitative summary of effect direction for oleic vs. linoleic acid across outcomes: insulin sensitivity, inflammation, lipid profile, oxidative stress, β cell function with color-coded by strength of evidence (preclinical only vs. human data).
Evidence Heatmap Summary for Oleic Acid (OA) and Linoleic Acid (LA) in T2DM
| Pathway / Outcome | OA (Oleic Acid) | LA (Linoleic Acid) |
|---|---|---|
| Insulin sensitivity | Dark green (+2) – strong and consistent evidence | Dark green (+2) – consistent support |
| Inflammation | Light green (+1) – moderate anti-inflammatory evidence | Gray (0) – mixed/insufficient evidence |
| Lipid metabolism | Dark green (+2) – strong cellular protection | Light green (+1) – moderate systemic benefits |
| Oxidative stress | Light green (+1) – moderate antioxidant protection | Gray (0) – limited evidence, prone to peroxidation |
| β-cell function | Dark green (+2) – strong cytoprotective effects | Light green (+1) – modest, context-dependent protection |
Color codes:
-
Dark green (+2): strong and consistent beneficial evidence
-
Light green (+1): moderate/mild beneficial evidence
-
Gray (0): mixed or insufficient evidence
-
Light red (-1): moderate/mild adverse evidence
-
Dark red (-2): strong and consistent adverse evidence
Discussion
The evidence reviewed highlights the multifaceted roles of oleic acid (OA, a MUFA) and linoleic acid (LA, a PUFA) in the management of type 2 diabetes mellitus (T2DM), underscoring their complementary contributions to metabolic regulation. Mechanistically, OA enhances insulin signaling by activating AMPK and Akt, preserving IRS1 function under lipotoxic conditions, and supporting β-cell insulin secretion, while counteracting the deleterious effects of saturated fatty acids such as palmitate 27. LA strengthens insulin action indirectly by inhibiting protein tyrosine phosphatase 1B (PTP1B), thereby amplifying the Akt and PI3K/AMPK pathways and protecting β-cells from lipotoxic stress 25. These complementary mechanisms highlight the potential of both fatty acids to preserve metabolic homeostasis under conditions of inflammation and nutrient excess 21,33.
Beyond glucose metabolism, OA demonstrates consistent anti-inflammatory and lipid-modulating properties 18. By displacing arachidonic acid in cellular membranes and suppressing cytokine-driven signaling pathways, OA reduces pro-inflammatory mediators while promoting an anti-inflammatory cytokine profile characterized by increased IL-10 and adiponectin levels 18. LA exerts more complex and context-dependent effects 31. While it can suppress adaptive immune responses by reducing Th1/Th17 differentiation and cytokine release, it may simultaneously enhance innate immune activation in neutrophils and macrophages 25,31,32. Human studies generally suggest that moderate LA intake does not exacerbate inflammation and may modestly improve adipose tissue function through PPAR-γ activation and increased adiponectin secretion 48. Thus, OA provides consistent anti-inflammatory protection 18, whereas LA modulates immune responses bidirectionally 50.
Both fatty acids contribute to improved lipid metabolism through distinct pathways 18,33. OA protects against intracellular lipotoxicity by channeling saturated fats into neutral triglyceride pools and enhancing mitochondrial β-oxidation, thereby preventing toxic lipid accumulation 34. LA improves systemic lipid profiles by lowering LDL cholesterol, upregulating hepatic LDL receptors, and shifting adipose tissue toward healthier PUFA-rich pools 35. Together, these actions reduce lipotoxic stress and support insulin sensitivity, further reinforcing their complementary metabolic roles 21.
In terms of oxidative stress, OA exerts strong antioxidant and cytoprotective effects, lowering reactive oxygen species (ROS) and reinforcing mitochondrial stability in β-cells, muscle, and endothelial cells 4,27. LA, being more prone to peroxidation, carries a theoretical risk of oxidative damage at very high intakes 36,37. However, human studies evaluating standard dietary levels do not show significant increases in oxidative biomarkers, and its insulin-sensitizing and lipid-modulating effects generally outweigh the potential risks 36,37. OA therefore provides more consistent direct antioxidant protection, while LA requires moderation to avoid peroxidative stress 57.
Limitations
Despite the breadth of the evidence reviewed, the absence of a systematic methodology remains a major limitation. Although multiple databases were searched to capture a wide range of studies, the review process was narrative rather than systematic. Without a reproducible search strategy, it is difficult to guarantee that all relevant studies were included, and it is possible that contradictory findings may have been overlooked. This limitation undermines both transparency and reproducibility, which are essential for scientific rigor, and thus must be explicitly acknowledged.
Another limitation pertains to the heterogeneity of dietary interventions and the potential for residual confounding in observational studies. Clinical trials and cohort studies often combine whole foods (such as olive oil or nuts), supplemented diets, and isolated fatty acids; however, these are not metabolically equivalent. This variability complicates data interpretation and makes it challenging to isolate the independent metabolic effects of oleic and linoleic acids. In observational studies, higher oleic acid intake frequently correlates with healthier dietary patterns, while linoleic acid intake may reflect a broader dietary composition. Even with statistical adjustments, disentangling these specific lipid effects from overall lifestyle factors remains difficult, leaving residual confounding as an unresolved issue.
Finally, limited geographic and ethnic diversity, along with the inability to assess publication bias, further constrain the evidence base. Most studies have been conducted in Western populations, with relatively little data derived from African, Asian, or Latin American cohorts. Given the genetic and cultural differences in dietary patterns, the generalizability of these findings across global populations remains uncertain. In addition, publication bias cannot be excluded, as positive findings are more likely to be published than null or negative results. Dose-dependent risks also require careful consideration: excessive oleic acid intake has been linked to adipogenesis 35, while excessive linoleic acid in high-sugar contexts may impair β-cell insulin secretion via endocannabinoid pathways 29,35. Although these risks have been identified, they remain insufficiently quantified. Collectively, these limitations highlight the need for long-term randomized controlled trials across diverse populations, utilizing systematic methodologies and explicit strategies to detect and mitigate publication bias.
Clinical Implications
The complementary actions of OA and LA underscore the importance of a balanced dietary intake in T2DM management. OA exerts consistent anti-inflammatory, antioxidant, and β-cell protective effects, while LA contributes to systemic lipid improvements and immune modulation 27,60. Together, these fatty acids enhance insulin sensitivity, reduce lipotoxic stress, and mitigate diabetes-related complications 18.
Clinically, substituting saturated fats with MUFA- and PUFA-rich foods represents a practical strategy to improve metabolic outcomes 18. Olive oil, nuts, seeds, and vegetable oils are accessible dietary sources that can be readily incorporated into daily diets 33. Moderate consumption of both OA and LA appears beneficial, whereas excessive intake may pose potential metabolic risks 57. Very high LA intake coupled with high-sugar diets has been linked to impaired β-cell insulin secretion, while excessive OA consumption may promote adipogenesis, thereby highlighting the need for dietary moderation 35,46.
The cardiometabolic benefits of these fatty acids are particularly relevant in T2DM, a condition characterized by a markedly elevated cardiovascular risk 18. The antioxidant properties of OA and the LDL-lowering effects of LA act synergistically to reduce oxidative stress and improve lipid profiles 21, offering dual protection against diabetes-related cardiovascular complications. Personalized nutrition approaches may further optimize clinical outcomes, as genetic, metabolic, and lifestyle factors significantly influence fatty acid metabolism and individual responsiveness 18.
Future research should prioritize long-term randomized controlled trials to establish optimal intake ranges, clarify dose-dependent effects, and explore the synergistic interactions between MUFAs and PUFAs, thereby refining dietary guidelines for individualized diabetes care.
Conclusion
Oleic acid (a MUFA) and linoleic acid (a PUFA) play complementary but distinct roles in the management of T2DM. OA enhances insulin signaling, reduces inflammation, protects β-cell viability, and provides antioxidant defenses, while LA improves lipid handling, lowers LDL cholesterol, and modulates immune responses in a context-dependent manner. Together, they mitigate lipotoxic stress, enhance insulin sensitivity, and reduce cardiometabolic risk, thereby underscoring their dietary relevance. Clinical evidence supports replacing saturated fats with MUFA- and PUFA-rich foods, such as olive oil, nuts, seeds, and vegetable oils. However, dose-dependent effects and methodological limitations must be considered, as excessive intake may pose metabolic risks. Personalized nutrition approaches may further optimize clinical outcomes. Future trials should define optimal intake ranges and clarify synergistic interactions, thereby supporting balanced consumption to improve overall metabolic health.
Abbreviations
AMPK: AMP-activated protein kinase; DAG: Diacylglycerol; ER: Endoplasmic reticulum; GLUT4: Glucose transporter type 4; GPR119: G protein-coupled receptor 119; HbA1c: Glycated hemoglobin; IDF: International Diabetes Federation; IL-1β: Interleukin-1 beta; IL-6: Interleukin-6; IL-10: Interleukin-10; IRS1: Insulin receptor substrate 1; JNK: c-Jun N-terminal kinase; LA: Linoleic acid; LDL: Low-density lipoprotein; MeSH: Medical Subject Headings; MUFA: Monounsaturated fatty acid; NF-κB: Nuclear factor kappa B; NLRP3: NOD-, LRR- and pyrin domain-containing protein 3; Nrf2: Nuclear factor erythroid 2-related factor 2; OA: Oleic acid; OEA: Oleoylethanolamide; PI3K: Phosphoinositide 3-kinase; PPAR-α: Peroxisome proliferator-activated receptor alpha; PPAR-γ: Peroxisome proliferator-activated receptor gamma; PTP1B: Protein tyrosine phosphatase 1B; PUFA: Polyunsaturated fatty acid; ROS: Reactive oxygen species; SFA: Saturated fatty acid; T2DM: Type 2 diabetes mellitus; Th1/Th17: T helper type 1 / T helper type 17; TLR4: Toll-like receptor 4; TNF-α: Tumor necrosis factor-alpha.
Acknowledgments
The author(s) thank colleagues and peers for their valuable input during manuscript preparation.
Author’s contributions
All authors equally contributed to this work, read and approved the final manuscript.
Funding
None.
Availability of data and materials
Not applicable.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Declaration of generative AI and AI-assisted technologies in the writing process
The authors declare that they have not used generative AI (a type of artificial intelligence technology that can produce various types of content including text, imagery, audio and synthetic data).
Competing interests
The authors declare that they have no competing interests.