

Direct reprogramming of somatic cells: an update
- Laboratory of Stem Cell Research and Application, University of Sciences, Vietnam National University, Ho Chi Minh City, Viet Nam
Abstract
Direct epigenetic reprogramming is a technique that converts a differentiated adult cell into another differentiated cell—such fibroblasts to cardiomyocytes—without passage through an undifferentiated pluripotent stage. This novel technology is opening doors in biological research and regenerative medicine. Some preliminary studies about direct reprogramming started in the 1980s when differentiated adult cells could be converted into other differentiated cells by overexpressing transcription-factor genes. These studies also showed that differentiated cells have plasticity. Direct reprogramming can be a powerful tool in biological research and regenerative medicine, especially the new frontier of personalized medicine. This review aims to summarize all direct reprogramming studies of somatic cells by master control genes as well as potential applications of these techniques in research and treatment of selected human diseases.
Cell fate and reprogramming
The human body originates from a totipotent stem cell, the zygote. Development and growth of an organism are due to proliferation and differentiation of these cells. Stem cell proliferation by self-renewal causes an increase in cell numbers, while stem cell differentiation causes an increase in cell types. Although all cells in the human body originate from a single cell, they play different roles. Their finalized specific functions are decided by mechanisms that are yet unclear, but it is considered that their functions are decided by their fates or programming (alterations in gene expression). From a single totipotent stem cell, generations of daughter cells are programmed into specific cell types that collaborate with each other to produce a completed body.
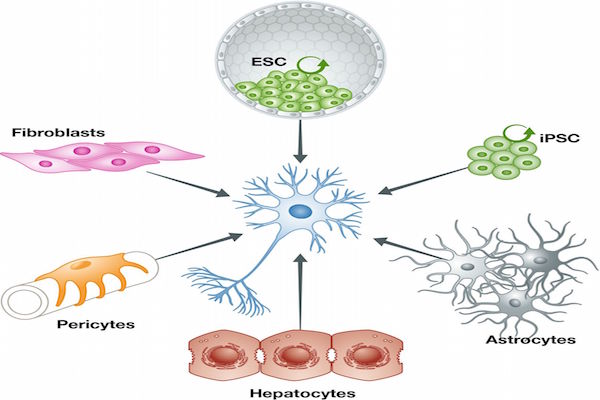
In the traditional view, cell fates cannot be modified, and stem cell differentiation is unidirectional, in which only uncommitted or undifferentiated cells can differentiate into committed or specific cells. However, to date, many studies prove that fully differentiated cells can reverse to pluripotent stem cells. This process is termed as“reprogramming” ( Figure 1 ).

Definitions of differentiation, reprogramming and direct reprogramming.
Reprogramming
The first attempt of the reprogramming technique was performed by Robert Briggs and Thomas King. In 1952, they injected an embryonic nucleus into an enucleated egg in the amphibian Rana pipiens (Briggs and King, 1952), advancing from an oocyte to the tadpole stage of development. However, this experiment failed when carried out with fully differentiated cells. These results made them conclude that differentiated nuclei cannot revert to a developing embryo King and Briggs, 1955. Conversely, John B. Gurdon successfully produced swimming tadpoles from transplantation of differentiated tadpole intestinal epithelial cell nuclei into enucleated eggs that were exposed to ultraviolet irradiation Gurdon, 1962. By this result, Gurdon concluded that differentiated somatic cell nuclei can revert to pluripotency.
Gurdon’s discovery opened a new field in animal development biology. He presented a new mechanism that changed the opinion of hundreds of biological scientists. This discovery was confirmed by Wilmut et al. in 1997. Similarly, for the first time in a mammal, Wilmut successfully created the sheep Dolly by injecting adult mammary epithelial cell nuclei into an enucleated sheep egg Wilmut et al., 1997. After these results, more than 10 different species, such as mouse, cow, pig, cat, and dog, have been “cloned” by the injection adult cell nuclei into oocytes, and the technique is also called “somatic cell nuclear transfer—SCNT.”
Although Gurdon showed that differentiated cell nuclei could be reprogrammed into the undifferentiated state to re-start development, many experiments also suggested that the intact, differentiated cells could be reprogrammed into undifferentiated cells. This was confirmed by Shinya Yamanaka in 2007. He choose 24 transcription factors related to embryonic stem cells, and from these 24 genes his group demonstrated that only four genes, Myc, Oct3/4, Sox2, and Klf4 could reprogram mouse embryonic fibroblasts into pluripotent stem cells Takahashi and Yamanaka, 2006. These stem cells were termed as induced pluripotent stem cells (iPSCs) by Yamanaka. iPSCs exhibit most of the characteristics of embryonic stem cells such as selfrenewal and long-term-multiple lineage differentiation, and have been especially useful in the production of mouse chimeras. In 2007, Yamanaka and James Thomson’s laboratories were the first to successfully produce human iPSCs Takahashi et al., 2007aYu et al., 2007. Yamanaka’s group used the four factors found in mouse: Oct4, Sox2, Klf4, and Myc (OSKM); Thomson used the set: Lin28, Nanog, Oct4, and Sox2. For these critical contributions in reprogramming technology, John Gurdon and Shinya Yamanaka shared a Nobel Prize in Physiology and Medicine in 2012.
Since 2006, iPSC technology has been continuously refined to produce iPSCs with higher efficiency and easier and safer production. In a study, OSKM was transfected in mouse embryonic fibroblasts by viral vectors Takahashi et al., 2007b. Four of these factors would activate the pluripotent status of differentiated cells Jaenisch and Young,2008. In human fibroblasts, Oct4 and Sox2, together with Nanog and LIN28, can reprogram them toward pluripotent cells Yu et al., 2007.
To improve the efficiency of reprogramming, subsequent studies used polycistronic vector containing four factors, chromatin-modifying chemicals, and mRNAs, in combination with activation or inhibition of various signaling pathways involved in the regulation of cell proliferation Chang et al., 2009Feng et al., 2009Heinrich and Dimmeler, 2012Kretsovali et al., 2012.
Some studies also significantly improved the safety of transgenes. In an early study, retroviral vectors were integrated into a genome, causing insertional mutagenesis. This technique is can be modified by utilizing non-integrating vectors Stadtfeld and Hochedlinger, 2010Stadtfeld et al., 2008. Further advances related to DNA-free transgenes using mRNAs or proteins were achieved Jia et al., 2010Warren et al.,2010Zhou et al., 2009.
With these improvements, clinical-grade iPSCs were developed in the recent years. Clinical grade iPSCs usually use donor cells such as fibroblasts, keratinocytes, and peripheral blood mononuclear cells (PBMCs), which are preferable for inducing pluripotency. Moreover, clinical-grade iPSCs need to be produced from safer techniques, reducing the likelihood of accidently creating tumor-forming cells.
Some safer techniques in gene transfection are used to produce vectors containing reprogramming genes. The first effort used F-deficient Sendai virus particles to induce pluripotency in somatic cells Dowey et al., 2012Fusaki et al., 2009. iPSCs produced using this method must be sub-cultured for 10–20 passages to remove the excess virus particles and to make virus-free iPSC lines. Later, an improvement in gene transfection using temperature-sensitive Sendai virus particles made it is easier to remove the virus particles by temperature shift (Ban et al., 2011).
Virus-free vectors carrying reprogramming factors have been studied since 2010 to replace the viral vectors. Episomal DNA can be used to transfect transgenes into adult cells. These virus-free vectors have important clinical applications because they are safer in manipulations as well as in the patients. There are two kinds of episomes: non-replicating episomal vectors and replicating episomal vectors. The iPSC production procedure using non-replicating episomal vectors is of low-yield; therefore, multiple transfections are suggested as a solution to increase the iPSC production efficacy Jia et al., 2010Okita et al., 2008. Improvements such as the use of minicircle or codonoptimized 4-in-1 minicircle (CoMiP) DNA vectors were devised Lu et al., 2013Okita et al., 2008.
Although DNA-based episome is considered safe to reprogram adult cells to iPSCs, in principle, foreign DNA can integrate into the host genome. Therefore, iPSCs must be screened to select free cells for further applications Gonzalez et al., 2009. To date, the safest technique of iPSC production is induction of pluripotency via mRNA Warren et al., 2012Yoshioka et al., 2013 or protein Kim et al., 2009Lee et al., 2012. These iPSCs are called “clean” iPSCs.
Together with improvement of iPSC production methods, some approaches using iPSCs in treatment were also developed. The most significant approach for clinical applications of iPSCs relates to the combination of iPSC technology and targeting editing of the iPSC genome. This combination helps to push iPSCs into clinical treatment, particularly for patients with genetic disorders. There are three ways to correct the mutated genes in iPSCs: the zinc finger nuclease (ZFN) system, the transcription activator-like effector nuclease (TALEN) system, and the clustered regularly interspaced short palindromic repeats (CRISPR) system Ding et al., 2013Hockemeyer et al., 2009Horii et al., 2013. By using these techniques, patient-specific iPSCs were successfully produced to treat epilepsy Parent and Anderson, 2015, myotonic dystrophy type 1 Xia et al., 2015, sickle erythrocytes Huang et al., 2015, retinal degenerative diseases Wiley et al., 2015, and recessive dystrophic epidermolysis bullosa Sebastiano et al., 2014.
Direct reprogramming
The direct reprogramming technique was discovered in the 1980s ( Table 1 ). In 1987, Davis et al. converted embryonic mouse fibroblasts into muscle cells by transfection of myogenic differentiation factor (MyoD) Davis et al., 1987. Similarly, MyoD was used to reprogram immature chondrocytes, smooth muscle cells, and retinal cells into muscle cells Choi et al., 1990. In the 1990s, some other transcription factors were discovered, particularly globin transcription factor 1 (Gata-1), that can reprogram avian monocyte precursors into erythrocytes, eosinophils, and megakaryocytes Kulessa et al., 1995.

Direct reprogramming presented in the Literature
Since 2000, several transcription factors were discovered and were successfully used to reprogram target cells such as pancreatic islet cells Zhou et al., 2008, neurons Fishman et al., 2015Vierbuchen et al., 2010, hepatocytes Huang et al., 2011Sekiya and Suzuki, 2011, endothelial cells Ginsberg et al., 2012Han et al., 2014, smooth muscle cells Karamariti et al., 2013, and hepatocyte like cells Simeonov and Uppal, 2014.
In recent years, in situ direct reprogramming as well as in vivo direct reprogramming has become important, as the ability to provide novel therapies is nearly in clinical applications. In vivo direct reprogramming is the usage of specific transcription factors to change target cell fate in the body without the need to isolate the target cells ( Table 1 ).
In early studies, it was shown that transcription factors can directly affect reprogramming. Recent studies indicated that there are at least five kinds of reprogramming factors that can directly reprogram adult cells into other phenotypic cells: transcription factors, epigenetic regulators, miRNAs, Small molecules, and pluripotency factors for direct reprogramming.
Transcription factors
Different from reprogramming techniques make adult cells pluripotent after receiving some key transcription factors causing epigenetic modifications, direct reprogramming mechanisms are still elusive. The most important mechanism is the effect of transcription factors that drive the phenotype changes in specific cells. By using transcription factors, transfected cells can change phenotype via activation of target genes. Interestingly, these changes can occur some hours after transfections Ieda et al., 2010, do not require cell division Heinrich et al., 2010Vierbuchen et al., 2010, and are stable after removal of reprogramming factors Huang et al., 2011Sekiya and Suzuki, 2011. Some authors have demonstrated that direct reprogramming of fibroblasts to neurons was hierarchical, established mechanisms dictate that fibroblasts gradually change with multiple steps to become neurons Wapinski et al., 2013.
Epigenetic regulators
Differentiated status of cells seems depend on epigenetic status of these cells. Transcription factors are known as important factors effecting to expression of lineage specific genes. However, gene expression also is effected by epigenetic regulators. In fact, there are three ways that epigenetic regulators effect gene expression. First, epigenetic regulators can decide the reprogramming process by themselves. For example, pancreatic beta cells can be reprogrammed into alpha cells by DNA methyltransferase Dnmt1 deficiency Dhawan et al., 2011. Second, epigenetic regulators can interact with exogenous factors to re-activate or suppress related gene expression. In the study by Takeuchi and Bruneau (2009), they showed that Baf60c – cardiac specific subunit of BAF chromatin remodeling complexes hold a particular role in the reprogramming from mouse mesoderm to cardiac myocytes that is helped by Gata4 – a transcription factor to bind to cardiac genes Takeuchi and Bruneau, 2009. Third, some epigenetic regulators act as epigenetic barriers that can prevent reprogramming. In fact, the inhibition or removal of histone deacetylases and polycomb repressor complex 2 (PRC2) can facilitate the reprogramming of germ cells into neurons Patel et al., 2012.
miRNAs
More and more studies proved that miRNAs play important roles in the reprogramming process. Some specific miRNAs such as miR-124, miR-9/9, miR-1, miR-133, miR-208, and miR-499 were demonstrated with reprogramming effects in fibroblasts. Overexpression of miR-9/9 and miR-124 in human fibroblasts can induce the expression of markers indicative of neuron-like cells Yoo et al., 2011. It seems that miRNAs can regulate some mechanisms relating to epigenetic reprogramming. In fact, miRNAs can directly stimulate or suppress target genes (Bartel, 2009) as well as regulate epigenetic regulators Neo et al., 2014. However, in general, miRNAs are not as efficient as transcription factors to induce epigenetic reprogramming.
Small molecules
Some small molecules were successfully used to produce iPSC Li et al., 2013b. The main advantage of small molecules is small structure, therefore they can more easily move across cellular membranes. By this advantage, small molecules are more richly investigated in recent studies. The biggest success in direct reprogramming by small molecules is the neural conversion process Kim et al., 2014Sayed et al., 2015. How the small molecules can reprogram the cell fate is a question that needs to be answered. In some cases, small molecules activate some pluripotency genes Hou et al., 2013 as well as transcription factors Yuan et al., 2013.
Pluripotency Factors for Indirect Reprogramming
Some pluripotency factors used to produce iPSC can directly reprogram some cell types such as cardiomyocytes Efe et al., 2011, neural stem cells or progenitors Wang et al., 2013, angioblast-like progenitor cells Kurian et al., 2013, endothelial cells Li et al., 2013a, pancreatic lineages Li et al., 2014, and hepatocytes Zhu et al., 2014. Ma et al. (2013) showed that pluripotent factors can reprogram adult cells into pluripotent cells with multiple steps and that at certain steps some cells’ fates are formed as transition stages of epigenetic reprogramming Ma et al., 2013. Moreover, overexpression of pluripotent factors can also induce differentiation Loh and Lim,2011.
Although direct reprogramming can produce the functional cells that can be used in translational applications as well as therapy, the main limitation of this technology is slow or non-proliferation of reprogrammed cells. Therefore, direct reprogramming should be improved in order to produce proliferating cells such as tissue specific stem cells or progenitor cells more than fully differentiated cells. In fact, some kinds of stem cells as well as progenitor cells were produced by direct reprogramming technology, including neural stem cells or progenitors Han et al., 2012Schindeler et al., 2015Thier et al., 2012, oligodendrocyte precursor cells Najm et al., 2013, hepatic stem cells Yu et al., 2013, HSCs Riddell et al., 2014, and hematopoietic multipotent progenitors (Batta et al., 2014)Sandler et al., 2014.
Invivo direct reproramming
As direct reprogramming technology is gradually perfected, especially its efficiency in combination with the tools of in situ gene therapy that were developed in previous studies. In vivo direct reprogramming has become more interesting as a novel therapy in regenerative medicine. Using in situ gene therapy strategies with direct reprogramming factors, some preclinical trials with a mouse model were successful in the conversion of various cerebral cell types into neurons Heinrich and Rouaux, 2015. By enhanced expression of Sox10 in Satellite Glial cells, Weider et al (2015) successfully induced these cells in vivo into oligodendrocyte- like cells Weider et al.,2015.
Particularly, reactive glial cells in the cortex of stabinjured or Alzheimer’s disease (AD) model mice can be directly reprogrammed into functional neurons in vivo using retroviral expression of a single neural transcription factor, NeuroD1 Guo et al., 2014. More importantly, cardiac injury model mice can be treated by in vivo direct reprogramming Jayawardena et al., 2015. miRNAs and lentiviral vectors were injected into these mice. After 5-6 weeks, cardiac function was improved, associated with existence of cardiac myocyte- like cells in injected sites.
Conclusion
Epigenetic reprogramming has seen rapid growth in recent years. Supported by some modern molecular biology techniques, reprogramming technology is becoming important and promising for wide use in basic research to translational research, and clinical application in the near future. Direct epigenetic reprogramming is a combination of stem cell therapy and gene therapy that can induce cell regeneration in an in situmanner. Many non-viral vectors and some novel reprogramming factors have facilitated direct reprogramming applications in preclinical models. Direct reprogramming, however, also faces with some challenges. Safety of vectors as well as technology must be investigated and carefully evaluated, especially in viral vector transfections or DNA transfection. Another challenge relates to control of reprogramming efficiency as well as specificity of target cells in vivo.
Abbreviations
AD: Alzheimer’s disease; PRC2: Polycomb repressor complex 2; ZFN: zinc finger nuclease; HSCs: Hematopoietic stem cells; iPSC: Induced pluripotent stem cells.